Полушария большого мозга | Компетентно о здоровье на iLive
Полушарие большого мозга
Полушарие большого мозга (hemispherium cerebralis) снаружи покрыто тонкой пластинкой серого вещества — корой большого мозга. Каждое полушарие имеет три поверхности: наиболее выпуклую верхнелатеральную (facies superolateral, hemispherii), плоскую, обращенную к соседнему полушарию медиальную (facies medialis hemispherii) и нижнюю (facies inferior hiispherii). Последняя имеет сложный рельеф, соответствующий внутреннему основанию черепа. Поверхности полушарий большого мозга отделены друг от друга краями: верхним (margo superior), нижнелатеральным (margo inferior) и нижнемедиальным (margo medialis). Наиболее выступающие кпереди и кзади участки полушария получили название полюсов: лобный полюс (polus frontalis), затылочный полюс (polus occipitalis) и височный полюс (polus tiporalis). Рельеф поверхностей полушарий очень сложен в связи с наличием более или менее глубоких борозд большого мозга и расположенных между ними валикообразных возвышений — извилин. Глубина, протяженность борозд и выпуклых извилин, их форма и направление очень изменчивы.
Верхнелатеральная поверхность полушария
В переднем отделе каждого полушария большого мозга находится лобная доля (lobus frontalis). Она заканчивается спереди лобным полюсом и ограничена снизу латеральной бороздой (sulcus lateralis; сильвиева борозда), а сзади глубокой центральной бороздой. Центральная борозда (sulcus centralis; роландова борозда) расположена во фронтальной плоскости. Она начинается в верхней части медиальной поверхности полушария большого мозга, рассекает поперек его верхний край, спускается, не прерываясь, по верхнелатеральной поверхности полушария вниз и заканчивается немного не доходя до латеральной борозды.
Лобные доли головного мозга
Кзади от центральной борозды находится теменная доля (lobus parietalis). Задней границей этой доли является теменно-затылочная борозда (sulcus parietooccipitalis). Эта борозда располагается на медиальной поверхности полушария большого мозга, глубоко рассекает верхний край полушария и переходит на его верхнелатеральную поверхность.
Теменная доля головного мозга
Затылочная доля (lobus occipitalis) располагается позади теменно-затылочной борозды и ее условного продолжения на верхнелатеральной поверхности полушария. По сравнению с другими долями она имеет небольшие размеры. Затылочная доля кончается затылочным полюсом (polus occipitalis). Борозды и извилины на верхнелатеральной поверхности затылочной доли очень изменчивы.
Затылочная доля головного мозга
Височная доля (lobus temporalis) занимает нижнебоковые отделы полушария и отделяется от лобной и теменной долей глубокой латеральной бороздой. Край височной доли, прикрывающий островковую долю, получил название височной покрышки (operculum temporale). Передняя часть височной доли образует височный полюс (polus temporalis). На боковой поверхности височной доли видны две борозды — верхняя и нижняя височные (sulci temporales superior et inferior), почти параллельные латеральной борозде. Извилины височной доли ориентированы вдоль борозд.
Височная доля головного мозга
Островковая доля, островок (lobus insularis, s. insula) находится в глубине латеральной борозды. Эту долю можно увидеть, если раздвинуть или удалить прикрывающие островок участки лобной, теменной и височной долей, которые получили наименование покрышки. Глубокая круговая борозда островка (sulcus circularis insulae) отделяет островок от окружающих его отделов мозга. На поверхности островка имеются извилины островка, длинная и короткие (gyri insulae, longus et breves). Между длинной извилиной, находящейся в задней части островка и ориентированной сверху вниз и вперед, и короткими извилинами, занимающими верхнепереднюю часть островка, находится центральная борозда островка (sulcuscentralis insulae). Нижнепередняя часть островка лишена борозд и имеет небольшое утолщение — порог островка (limen insulae).
Медиальная поверхность полушария
Все доли полушария, за исключением островковой, принимают участие в образовании его медиальной поверхности. Над мозолистым телом, отделяя его от остальных отделов полушария, находится борозда мозолистого тела (sulcus corporis callosi). Огибая сзади валик (splenium) мозолистого тела, эта борозда направляется книзу и вперед и продолжается в борозду гиппокампа, или гиппокампальную борозду (sulcus hippocampi, s. hippocampalis). Выше борозды мозолистого тела находится поясная борозда (sulcus cinguli). Эта борозда начинается кпереди и книзу от клюва мозолистого тела, поднимается вверх, затем поворачивается назад и следует параллельно борозде мозолистого тела. Заканчивается борозда выше и кзади от валика мозолистого тела как подтеменная борозда (sulcus subparietalis). На уровне валика мозолистого тела от поясной борозды вверх ответвляется краевая часть (pars marginalis, BNA), уходящая вверх и кзади к верхнему краю полушария большого мозга. Между бороздой мозолистого тела и поясной бороздой находится поясная извилина (gyrus cinguli), охватывающая мозолистое тело спереди, сверху и сзади. Сзади и книзу от валика мозолистого тела поясная извилина суживается, образуя перешеек поясной извилины (isthmus gyri cinguli). Далее книзу и кпереди перешеек переходит в более широкую извилину гиппокампа, или парагиппокампальную извилину (gyrus parahippocampalis), ограниченную сверху бороздой гиппокампа. Поясная извилина, перешеек и парагиппокампаль-ная извилина известны как сводчатая извилина (gyrus fornicatus — BNA). В глубине борозды гиппокампа находится довольно тонкая полоска серого цвета, разделенная мелкими поперечными бороздками, — зубчатая извилина (gyrus dentatus). Участок медиальной поверхности полушария, находящийся между поясной бороздой и верхним краем полушария, относится к лобной и теменной долям.
Кпереди от верхнего края центральной борозды находится медиальная поверхность верхней лобной извилины, а непосредственно к указанному участку центральной борозды прилежит парацентральная долька (lobulus paracentralis), ограниченная сзади краевой частью поясной борозды. Между краевой частью спереди и теменно-затылочной бороздой сзади находит предклинье (precuneus) — принадлежащий теменной доле участок полушария большого мозга.
На медиальной поверхности затылочной доли расположены сливающиеся друг с другом под острым углом, открытым кзади, две глубокие борозды: теменно-затылочная, отделяюшая теменную долю от затылочной, и шпорная борозда (sulcus calcaneus). Последняя начинается на медиальной поверхности затылочного полюса и направляется вперед до перешейка поясной извилины. Участок затылочной доли, лежащий между теменно-затылочной и шпорной бороздами и имеющий форму треугольника, обращенного вершиной к месту слияния этих борозд, называется клином (cuneus). Хорошо заметная на медиальной поверхности полушария шпорная борозда ограничивает сверху язычную извилину (gyrus hingualis), простирающуюся от затылочного полюса сзади до нижней части перешейка поясной извилины cнизу от язычной извилины располагается коллатеральная борозда (sulcus collateralis), принадлежащая уже нижней поверхности полушария.
Нижняя поверхность полушария
Рельеф нижней поверхности полушария очень сложен. Передние отделы этой поверхности образованы лобной долей полушария, позади которой выступает височный полюс, а также находятся нижние поверхности височной и затылочной долей, переходящие одна в другую без заметных границ.
На нижней поверхности лобной доли, несколько латеральнее и параллельно продольной щели большого мозга, проходит обонятельная борозда (sulcus olfactorius). Снизу к ней прилежат обонятельная луковица и обонятельный тракт, переходящий сзади в обонятельный треугольник. В области этого треугольника видны медиальная и латеральная обонятельные полоски (striae olfactoriae medialis et lateralis). Участок лобной доли между продольной щелью большого мозга и обонятельной бороздой получил название прямой извилины (gyrus rectus). Поверхность лобной доли, лежащая латеральнее обонятельной борозды, разделена неглубокими глазничными бороздами (sulci orbitales) на несколько изменчивых по форме, расположению и размерам глазничных извилин (gyri orbitales).
В заднем отделе нижней поверхности полушария хорошо различима коллатеральная борозда, расположенная книзу и латерально от язычной извилины на нижней поверхности затылочной и височной долей, латеральнее парагиппокампальной извилины. Несколько кпереди от переднего конца коллатеральной борозды находится носовая борозда (sulcus rhinalis). Она ограничивает с латеральной стороны изогнутый конец парагиппокампальной извилины — крючок (lincus). Латеральнее коллатеральной борозды лежит медиальная затылочно-височная извилина (gyrus occipitotemporalis medialis). Между этой извилиной и расположенной кнаружи от нее латеральной затылочно-височной извилиной (gyrus occipitotemporalis lateralis) находится затылочно-височная борозда (sulcus occipitotemporalis). Границей между латеральной затылочно-височной и нижней височной извилинами служит не борозда, а нижнелатеральный край полушария большого мозга.
Ряд отделов головного мозга, расположенных преимущественно на медиальной поверхности полушария и являющихся субстратом для формирования таких общих состояний, как бодрствование, сон, эмоции, мотивации поведения и др., выделяют под названием лимбическая система. Эти реакции сформировались в связи с первичными функциями обоняния (в филогенезе), поэтому их морфологической основой являются отделы мозга, которые развиваются из нижнелатеральных отделов мозгового пузыря и относятся к так называемому обонятельному мозгу (rhinencephalon). Лимбическую систему составляют обонятельная луковица, обонятельный тракт, обонятельный треугольник, переднее продырявленное вещество, расположенное на нижней поверхности лобной доли (периферический отдел обонятельного мозга), а также поясная и парагиппокампальная (вместе с крючком) извилины, зубчатая извилина, гиппокамп (центральный отдел обонятельною мозга) и некоторые другие структуры. Включение этих отделов мозга в лимбическую систему оказалось возможным в связи с общими чертами их строения (и происхождения), наличием взаимных связей и сходством функциональных реакций.
Полушария большого мозга
Большой мозг (cerebrum) представляет собой наиболее массивный отдел головного мозга и занимает большую часть полости мозгового черепа Продольная щель большого мозга (fissura Топ gitudinahs cerebn) делит большой мозг на два полушария (hemisphenum cerebn dextrum et sinistrum).
Поверхность полушарий покрыта слоем серого вещества — корой большого мозга — наиболее поздним по развитию и наиболее совершенным отделом нервной системы. Большое количество мозговых извилин(gyri), отделенных одна от другойбороздами (suici), значительно увеличивают площадь полушарий, что принципиально отличает мозг человека от мозга других млекопитающих. Наиболее глубокие борозды делят каждое полушарие на
Самой большой и глубокой бороздой верхнелатеральной поверхности большого мозга (. 17, а) является латеральная борозда(sul. lateralis), в глубине которой залегает островковая доля (insula). Покрывающие его части соседних долей называются покрышкой (operculum). Латеральная борозда ограничивает сверху височную долю, отделяя ее от лобной и теменной.
Вторая большая борозда верхнелатеральной поверхности, центральная(sul. centralis), отделяет лобную долю от теменной. Границей между затылочной и теменной долями служит теменно-затылочная борозда (sul. parietooccipitalis), располагающаяся преимущественно на медиальной поверхности и только немного заходящая на верхнелатеральную.
Впереди центральной борозды параллельно ей проходит пред-центральная борозда(sul. precentralis). Извилина, расположенная между центральной и предцентральной бороздами, называется предцентральной (gyrus precentralis). В лобной доле различают три идущие горизонтально извилины (gyri frontales superior, me-dius et inferior), разграниченные лобными бороздами (sul. frontales superior et inferior).
Позади центральной борозды располагается постцентральная борозда (sul. postcentralis), между ними находится постцентральная извилина (gyrus postcentralis). Поперечно расположенной внутритеменной бороздой (sul. intraparietalis) теменная доля делится на верхнюю и нижнюю теменные дольки (lobuli parietal is superior et inferior). В последней различают надкраевую извилину (gyrus supramarginalis), огибающую боковую борозду, и угловую (gyrus angularis), огибающую верхнюю височную борозду.
На верхнелатеральной поверхности височной доли проходят две горизонтальные борозды (sull. temporales superior et inferior), которые разделяют продольно расположенные верхнюю, среднюю и нижнюю височные извилины (gyrus temporales superior medius et inferior).
Борозды верхнелатеральной поверхности затылочной доли крайне непостоянны по количеству и направлению.
На медиальной поверхности полушария (. 17,6) дугообразно над мозолистым телом проходит борозда мозолистого тела (sul. corporis callosi) и параллельно ей—поясная борозда (sul. cinguli [cingulatus]). Эти борозды ограничивают поясную извилину(gyrus cinguli [cingulatus]. Последняя огибает колено мозолистого тела и переходит в подмозолистое поле (area subcal-losa). Кзади поясная извилина, обогнув мозолистое тело, образует перешеек поясной извилины (isthmus gyri cinguli) и перехо
дит в парагиппокампальную извилину(gyrus parahyppocampalis), или извилину гиппокампа, располагающуюся вдоль внутренней поверхности височной доли. Эта извилина отграничена от мозгового ствола бороздой гиппокампа (sul. hyppocampi). Ее нижней границей является коллатеральная борозда (sul. collateralis). Вентральный конец парагиппокампальной извилины загнут кверху и кзади, образуя крючок (uncus). В глубине борозды морского конька располагается узкая зубчатая извилина (gyrus dentatus).
Помимо описанных извилин на медиальную поверхность полушарий переходят извилины лобной, теменной и затылочной долей. На границе между лобной и теменной долями располагается пара-центральная долька (lobulus paracentralis), которая представляет собой переход предцентральной извилины лобной доли в зацент-ральную извилину теменной доли.
В задней части медиальной поверхности выделяют две глубокие борозды — теменно-затылочную и шпорную (sull. parietooccipitalis et calcarinus). Над шпорной бороздой лежит извилина— клин (cuneus), ниже—язычная извилина (gyrus lingualis). Впереди клина, между теменно-затылочной бороздой и поясной бороздой, располагается предклинье (precuneus) — четырехугольная извилина, относящаяся к теменной доле.
Борозды и извилины нижней поверхности полушарий (основания) головного мозга (. 17,б) относятся к различным долям. На нижней поверхности лобной доли различают обонятельную борозду(sul. olfactorius), занятуюобонятельной луковицей(bul-bus olfactorius) иобонятельным трактом(tr. olfactorius) и ограничивающую сбоку прямую извилину (gyrus rectus). Границу между лобной и височной долями образует здесь выходящая на нижнюю поверхность головного мозга латеральная борозда. На нижней поверхности височной доли простирается нижняя височная борозда. Ряд других борозд и извилин переходит сюда с верхнелатеральной и медиальной поверхностей полушарий.
Толщина коры большого мозгасоставляет 1,3—4,5 мм, лишь в парацентральной дольке достигает 10 мм, т. е. не везде одинакова, что связано с различным функциональным значением ее отдельных областей — в соответствии с биогенетическим законом соответствия морфологической эволюции эволюции функциональной.
Микроскопически кора большого мозга состоит из ряда клеток и волокон. Особенности миелиновых нервных волокон в различных участках коры изучает миелоархитектоника.В основу современной цитоархитектоники, учения об особенностях количества, формы, расположения клеток коры, легли работы киевского анатома В. А. Беца, который в 1874 г. описал гигантские клетки (клетки Беца) в предцентральной области коры большого мозга человека и животных, а в дальнейшем изучил архитектонику (он же и ввел этот термин) многих других областей коры.
В основном в коре различают шесть слоев:
1) молекулярный, находящийся непосредственно под мягкой мозговой оболочкой и характеризующийся бедностью клеточных элементов;
2) наружный зернистый, состоящий из большого количества мелких зернистых клеток;
3) слой малых и средних пирамидных клеток, величина и плотность которых варьируют в разных областях;
4) внутренний зернистый, состоящий из густо расположенных мелких клеток, чем напоминает наружный зернистый слой;
5) ганглионарный, содержащий большие пирамидные клетки (Беца), которые дают начало пирамидным путям;
6) слой полиморфных клеток треугольной и веретенообразной формы, непосредственно прилегающий к белому веществу.
Указанный тип строения коры имеет много вариаций, так как в различных ее участках слои получают разное развитие. В связи с цитоархитектоническими особенностями кору большого мозга принято делить на ряд областей,в каждой из которых различаютполя,имеющие порядковое обозначение. Наибольшее распространение получилацитоархитектоническая карта Бродмана,который различает 11 областей и 52 поля.
Белое веществополушарий занимает пространство между корой и базальными ядрами (полосатое тело, хвостатое и чечевице-образное ядра — ограда — claustrum, миндалевидное тело — corpus amygdaloideum, особенно тесно связанное лимбическими структурами мозга). Его массу составляют нервные волокна, идущие в разных направлениях и образующие проводящие пути. Нервные волокна делятся на проекционные, ассоциативные и комиссу-ральные.
Проекционные волокнасвязывают кору большого мозга со всеми нижележащими отделами центральной нервной системы и в белом веществе полушарий образуютлучистый венец(corona ra-diata). Они могут быть нисходящими (центробежными) (trr. cor-ticospinales, tr. corticonuclearis) и восходящими (центростремительными) (tr. thalamocorticalis).
Ассоциативные волокна,короткие и длинные, соединяют между собой различные участки в пределах одного и того же полушария (fibrae arcuatae cerebri, fasc. cinguli, fascc. longitudinales superior et inferior и др.).
Комиссуральные волокнавходят в состав спаек, соединяющих симметричные участки коры обоих полушарий. Как уже отмечалось, самой большой спайкой является мозолистое тело (corpus callosum) — филогенетически более молодое образование. Две другие мозговые спайки (comissura anterior, comissura fornicis), соединяющие между собой древние области коры (обонятельные доли, парагиппокампальные извилины), относятся к филогенетически древним образованиям.
Учение о локализации функций в коре большого мозгаразвивалось во взаимодействии двух противоположных концепций — анти-локализационизма, или эквипонтециализма (Флуранс, Лешли), отрицающего локализованность функций в коре, и узкого локали-зационного психоморфологизма, пытавшегося в своих крайних вариантах (Галль) локализовать в ограниченных участках мозга даже такие психические качества, как честность, скрытность, любовь к родителям и т. д. Большое значение имело открытие
фритчем и Гитцигом в 1870 г. участков коры, раздражение которых вызывало двигательный эффект. Другими исследователями также были описаны области коры, связанные с кожной чувствительностью, зрением, слухом. Клиницисты-неврологи и психиатры свидетельствуют также о нарушении сложных психических процессов при очаговых поражениях мозга. Брока показал, что поражение задней трети нижней лобной извилины левого полушария вызывает моторную афазию. В е р н и к е описал сенсорную афазию при патологических процессах в задней трети верхней височной извилины. Большой интерес вызвало сообщение Пенфилда о том, что электрическое раздражение коры височной доли у больных с птупами височной эпилепсии может вызывать воспоминания о прошедшем, которые внезапно оканчиваются, как только электрод удаляют (причем повторное раздражение той же точки спустя некоторое время может вызывать то же воспоминание). Так подкрепились представления о наличии закономерных зависимостей расстройства функции от локализации патологического очага.
Концепция локализационизма в свое время была прогрессивной по сравнению со взглядами эквипотенциоалистов, однако в дальнейшем она стала тормозить развитие медицинской науки, требующей четкого разграничения локализации симптомов и функций. Более сложные и филогенетически более молодые функции не могут быть узко локализованы, в их осуществлении участвуют обширные участки коры большого мозга или даже весь мозг в целом.
Основы современного принципиально нового взгляда на локализацию функций в головном мозге были заложены И. П. Павловым в его учении об анализаторахиучении о динамической локализации функций.По И. П. Павлову,анализатор —это сложный, функционально единый нейронный ансамбль, служащий для разложения (анализа) внешних или внутренних раздражителей на отдельные элементы. Он начинается рецептором на периферии и оканчивается в коре большого мозга. Корковые центры являются корковыми отделами анализаторов. И. П. Павлов внес существенные изменения в прежнее понятие об ограниченности территорий корковых центров. Он показал, что корковое представительство не ограничивается зоной проекции соответствующих проводников, далеко выходя за ее пределы, и что корковые зоны различных анализаторов перекрывают друг друга. Этот вывод И. П. Павлова вполне согласуется с новейшими нейрофизиологическими данными.
Итогом исследований И. П. Павлова явилось учение о динамической локализации функций, предполагающее возможность участия одних и тех же нервных структур в обеспечении различных функций. Под локализацией функций стали подразумевать формирование сложных динамических структуриликомбинационных центров,состоящих из мозаики возбужденных и заторможенных далеко отстоящих пунктов нервной системы, объединенных в общей работе в соответствии с характером необходимого конечного результата.
Свое дальнейшее развитие учение о динамической локализации функций получило в трудах П. К. Анохина, создавшего концепцию функциональной системыкак круга определенных физиологических проявлений, связанных с выполнением какой-либо определенной функции. Функциональная система включает каждый раз в разных сочетаниях различные центральные и периферические структуры: корковые и глубинные нервные центры, проводящие пути, периферические нервы, исполнительные органы. Одни и те же структуры могут входить во множество функциональных систем, в чем и выражается динамичность локализации функций.
Представление о нервном центре предполагает расположение нервных клеток, преимущественно обеспечивающих определенную функцию, в конкретных зонах коры большого мозга. В зависимости от отношения различных участков коры к другим образованиям мозга Г. И. Поляков выделяет три типа корковых полей. Первичные поля(ядра анализаторов, по И. П. Павлову) соответствуют тем архитектоническим полям, где заканчиваются чувствительные проводящие пути; это проекционные зоны, например ядро анализатора общей чувствительности в постцентральной извилине, ядро зрительного анализатора по краям и в глубине шпор-ной борозды.Вторичные поля(периферические отделы ядер анализаторов, по И. П Павлову) расположены вблизи первичных. В этих зонах, непосредственно не связанных с проводящими путями, происходит более детальная обработка информации, поступающей в первичные поля.Третичные полязанимают остальную, большую часть коры. Это ассоциативные зоны, располагающиеся в местах перекрытия отдельных анализаторов.
Третичные поля, не связанные непосредственно с периферией или с нижележащими отделами нервной системы, имеют связи с другими участками коры, в том числе и с проекционными зонами. Примером ассоциативной зоны может быть участок теменной доли, являющийся центром стереогноза.Поступающие в постцентральную извилину раздражения идут по таламокорковым путям — в результате этого возникают элементарные ощущения, испытываемые при ощупывании предмета (тепла, холода, тактильные, формы, величины и др.). По ассоциативным волокнам эти ощущения передаются в центр стереогноза, где благодаря его обширным связям с другими анализаторами создается образ предмета.
Кора большого мозга у человека характеризуется значительным развитием ассоциативных зон,которые в несколько раз превышают аналогичные зоны даже у высших животных.
В настоящее время в коре выделяют интегративно-пусковые зоны,функции которых заключаются в интеграции различных афферентных воздействий и формировании на этой основе пусковой команды, обеспечивающей ответную реакцию организма. Примером может служить двигательная зона.
Большинство корковых центров располагается в коре симметрично, однако имеются и непарные центры.Это, главным образом, центры, обеспечивающие речь и связанные с нею функции (чтение, счет, письмо). Эти зоны коры располагаются у праворуких в левом полушарии мозга. Правому полушарию также свойственны определенные специфические функции. Доказано преимущественное его значение в осуществлении ряда высших функций коры — восприятии пространства, времени, музыки и др.
Специфический для человека процесс познания осуществляется при участии обоих полушарий мозга, причем начальные ступени познания, или этап созерцания (переработка различной информации, создание образов окружающих объектов), связаны в большей степени с правым полушарием, а высшие ступени (этап абстрактного мышления) — с левым, связанным с мозговым обеспечением деятельности второй сигнальной системы.
На медиальной поверхности полушарий большого мозга, ближе к нижней их поверхности, расположены филогенетически древние лимбические структуры,которые остаются относительно неизменными в ходе эволюции и получили свое название за их расположение вокруг места прикрепления полушарий головного мозга к переднему концу мозгового ствола. Лимбическую зону впервые описал Брока (большая лимбическая доля Брока), но Папе ц впервые высказал мысль, что поясная извилина, гиппокамп и их связи, особенно связи с гипоталамусом, являютсяморфологическим субстратом эмоций.Он предположил, что афферентные импульсы на уровне таламуса делятся на “поток движения”, “поток мысли” и “поток чувства”. Последний, согласно его концепции, из таламуса направляется в гипоталамус и циркулирует по следующему кругу(круг Папеца):сосцевидные тела—сосцевидно-таламический пучок — передние ядра таламуса — поясная извилина — парагиппокампальная извилина — свод — сосцевидные тела. Гипотеза Папеца явилась фундаментом для современных представлений о лимбической и ретикуло-гипоталамо-лимбической системах. Благодаря связям с гипоталамусом и ретикулярной формациейлимбическая система(син.— обонятельный мозг (rhi-nencephalon), висцеральный мозг по Мак—Лину, эмоциональный мозг по Конорскому) имеет непосредственное отношение к поддержанию тонуса корковых нейронов, к нервным механизмам регуляции сна и бодрствования, координации вегетативно-эндокринных функций организма. Лимбическая система играет существенную роль в определении характера опосредуемых через кору ответных реакций на раздражения, в реализации положительных и отрицательных влияний на психическую деятельность, в механизмах памяти, в формировании нервных и эндокринных коррелятов эмоций, врожденного и эмоционального поведения, темперамента.
Большой мозг.
План лекции.
Большой мозг и его строение.
Особенности строения коры большого мозга и методы изучения функций коры.
Локализация функций в коре большого мозга.
Базальные ядра, лимбическая система и функции этих образований.
Биоэлектрическая активность головного мозга и методы ее изучения.
ЦЕЛЬ: Знать топографию и строение большого мозга: коры, базальных подкорковых ядер, лимбической системы.
Представлять локализацию функций в коре большого мозга, функции базальных ядер и лимбической системы, основные типы ритмов электроэнцефалограммы.
Уметь показывать на плакатах, муляжах и планшетах доли полушарий большого мозга, мозолистое тело, базальные ядра, боковые желудочки.
Большой мозг (cerebrum), или конечный мозг (teiencephalon), развивается из переднего (первого) мозгового пузыря. В процессе эволюции большой мозг возник позднее других отделов головного мозга, но достиг у человека наивысшего развития. По своей массе и величине он значительно превосходит все другие отделы головного мозга. Большой мозг состоит из двух полушарий — левого и правого, разделенных продольной щелью и соединяющихся между собой в глубине этой щели при помощи мозолистого тела, передней и задней спаек, а также спайки свода. Полости большого мозга образуют левый (первый) и правый (второй) боковые желудочки. Каждое полушарие большого мозга состоит из наружных покровов — коры (плаща), глубжележащего белого вещества и расположенных в нем скоплений серого вещества (базальных ядер). С полушариями большого мозга сращены таламусы и ножки мозга. Граница между большим и следующим за ним промежуточным мозгом проходит в том месте, где внутренняя капсула прилежит к латеральной стороне таламуса. Между полушариями и мозжечком имеется поперечная щель большого мозга.
На каждом полушарии различают 3 поверхности: верхнелатеральную — выпуклую, медиальную — плоскую и нижнюю — неровную, лежащую на основании черепа. Наиболее выступающие кпереди и кзади участки полушария получили названия полюсов: лобный полюс, затылочный полюс и височный полюс. Поверхности полушарий испещрены извилинами и бороздами. Извилины представляют собой валики (возвышения) мозгового вещества, а борозды — углубления между извилинами. Наличие борозд увеличивает поверхность коры полушарий большого мозга без увеличения его объема. В каждом полушарии различают 5 долей: лобную, теменную, височную, затылочную и островковую (островок И.Рейля).
Лобная доля занимает передний отдел полости черепа, включая переднюю черепную ямку, и отграничена от находящейся позади нее теменной доли центральной, или роландовой, бороздой.
Теменная доля находится сзади центральной борозды.
Височная доля расположена в средней черепной ямке и отделена от лобной и теменной долей глубокой латеральной (сильвиевой) бороздой.
Затылочная доля лежит над мозжечком в заднем отделе полости черепа. Между ней и теменной долей на медиальной поверхности полушария проходит теменно-затылочная борозда.
Островковая доля находится в глубине латеральной борозды. Ее можно увидеть, если раздвинуть или удалить прикрывающие островок участки лобной, теменной и височной долей, которые получили наименование покрышки.
Медиальная поверхность полушария имеет две концентрически расположенные извилины. Одна из них находится над мозолистым телом выше борозды мозолистого тела и называется поясной извилиной. Сзади и книзу от мозолистого тела поясная извилина суживается, образуя перешеек поясной извилины. Далее внизу и кпереди перешеек переходит во вторую более широкую извилину гиппокампа, или парагиппокампальнук извилину, ограниченную сверху бороздой гиппокампа. Поясная извилина перешеек и парагиппокампальная извилина образуют вместе сводчатую извилину. Передний изогнутый конец парагиппокампальной извилины называется крючком. Парагиппокампальная извилина отделяет височную долю от ствола мозга. Сводчатая извилина относится к лимбической (лат. limbus — кромка, кайма) области, входящей в состав лимбической системы.
Большой мозг построен из серого и белого вещества. Серое вещество снаружи полушария образует плащ, или кору большого мозга, в глубине полушария — подкорковые (базальные) ядра. Между корой и подкорковыми ядрами располагается белое вещество.
Внутри каждого полушария имеется полость, называемая боковым желудочком. В каждом желудочке различают центральную часть (в глубине теменной доли), от которой отходят 3 рога: передний (лобный), задний (затылочный) и нижний (височный). В центральной части и нижнем роге находится сосудистое сплетение бокового желудочка, продуцирующее спинномозговую жидкость.
Кора большого мозга — высший отдел ЦНС, формирующий деятельность организма как единого целого в его взаимоотношениях с окружающей средой. Она является наиболее молодым образованием ЦНС. С появлением коры происходит кортиколизация функций, т.е. регуляция функций организма перемещается из нижних отделов ЦНС в кору. Кора начинает контролировать все процессы, протекающие в организме, а также всю деятельность человека. По И.П.Павлову, кора является распорядителем и распределителем всех функций и всей деятельности организма. Кора — это вместилище всей нашей интеллектуальной жизни, это мастерская наших желаний, мыслей, воли и чувств. Деятельность коры большого мозга вместе с ближайшими подкорковыми ядрами носит название высшей нервной деятельности (ВНД).
Кора большого мозга представляет собой слой серого вещества толщиной от 1,5 до 5 мм. За счет большого количества складок площадь коры большого мозга составляет около 2200-2500 см2 (0,2-0,25 м2). В коре содержится от 14 до 17 млрд. нейронов, большая часть из которых (90%) сгруппирована в шесть слоев (пластинок) и образуют неокортекс (новую кору) – высший отдел соматической нервной системы. Из этих шести слоев нижние (V и VI слои) являются преимущественно началом эфферентных путей; в частности V слой состоит из пирамидных клеток, аксоны которых составляют пирамидную систему. Средние слои (III и IV слои) связаны преимущественно с афферентными путями, а верхние (I и II слои) относятся к ассоциативным нейронам и ассоциативным путям коры. Шестислойный тип коры видоизменяегся в различных областях как в смысле толщины и расположения слоев, так и состава клеток.
У человека неокортекс (новая кора) занимает 95,6% площади всей коры большого мозга. Остальную часть коры занимает другой отдел — палеокортекс (древняя кора — греч. palaios — древний). В отличие от неокортекса эта часть коры обладает более простой трехслойной структурой. Процессы, протекающие в палеокортексе, не всегда отражаются в сознании. К палеокортексу относят филогенетически самые древние и небольшие отделы коры, входящие в состав лимбической системы («обонятельного мозга»). Однако рассматривать эти отделы только как корковый отдел обонятельного анализатора нельзя, так как здесь расположены высшие корковые вегетативные центры.
Все пространство между серым веществом коры большого мозга и базальными ядрами занято белым веществом. Оно состоит из большого количества нервных волокон, идущих в различных направлениях и образующих проводящие пути конечного мозга. Эти нервные волокна могут быть трех видов:
ассоциативные (короткие или длинные), соединяющие между собой различные участки одного и того же полушария;
комиссуральные, связывающие чаще всего одинаковые симметричные участки двух полушарий; самая большая мозговая спайка — мозолистое тело связывает между собой части обоих полушарий;
проекционные (проводящие) волокна, осуществляющие связь с другими отделами ЦНС до спинного мозга включительно. Они обычно длинные, проводят возбуждение центростремительно, по направлению к коре, а другие волокна, наоборот, — центробежно, т.е. от коры.
 Для
изучения функций коры применяют следующие
методы:
Для
изучения функций коры применяют следующие
методы:
экстирпация, т.е. оперативное удаление отдельных участков коры;
метод электрического, химического и температурного раздражений различных зон коры;
метод электроэнцефалографии, т.е. регистрации биопотенциалов мозга, метод вызванных потенциалов;
метод условных рефлексов, разработанный И.П.Павловым;
 клинический
метод — изучение деятельности отдельных
органов и систем
при повреждении коры (кровоизлияние,
ранение, опухоль и т.д.).
клинический
метод — изучение деятельности отдельных
органов и систем
при повреждении коры (кровоизлияние,
ранение, опухоль и т.д.).
 клинический
метод — изучение деятельности отдельных
органов и систем
при повреждении коры (кровоизлияние,
ранение, опухоль и т.д.).
клинический
метод — изучение деятельности отдельных
органов и систем
при повреждении коры (кровоизлияние,
ранение, опухоль и т.д.).Роль отдельных областей коры большого мозга впервые была изучена в 1870 г. немецкими учеными Г.Фричем и Е.Гитцигом. Установлено, что разные участки коры ведают определенными функциями. Было создано учение о локализации функций в коре большого мозга. Отечественными авторами в это учение было внесено много новых данных. Так, например, киевский анатом В.А.Бец в 1874 г. доказал, что каждый участок коры отличается по строению от других участков мозга. Этим было положено начало учению о разнокачественности коры головного мозга. И.П.Павлов рассматривал кору полушарий большого мозга как сплошную воспринимающую поверхность, как совокупность корковых концов анализаторов. Он доказал, что корковый конец анализатора — это не какая-либо строго очерченная зона. В коре большого мозга различают ядро и рассеянные элементы. Ядро — это место концентрации нейронов коры, составляющих точную проекцию всех элементов определенного рецептора, где происходит высший анализ, синтез и интеграция функций. Рассеянные элементы могут располагаться как по периферии ядра, так и на значительном расстоянии от него. В них совершаются более простые анализ и синтез. Наличие рассеянных элементов при разрушении (повреждении) ядра отчасти позволяет компенсировать нарушенную функцию.
По наиболее распространенной классификации К.Бродмана в коре выделено 52 клеточных поля, каждое из которых имеет свой порядковый номер (1,2,3…52).
В зависимости от функциональных особенностей в коре выделяют моторные (двигательные), сенсорные (чувствительные) и ассоциативные зоны, осуществляющие связи между различными зонами коры. Нейтральные (немые) зоны в коре, как правило, отсутствуют.
Наиболее важные функциональные зоны коры.
Моторные зоны.
1) Моторная (двигательная) зона коры представлена в передней центральной (предцентральной) извилине лобной доли и парацентральной дольке. При неполном повреждении предцентральной извилины наблюдаются парезы (ослабление движений) скелетной мускулатуры на противоположной стороне, при полном повреждении — параличи (отсутствие движений), а при раздражении — разнообразные сокращения скелетных мышц.
Сенсорные зоны.
2) Зона кожной чувствительности (тактильной, болевой и температурной) представлена в задней центральной (постцентральной) извилине теменной доли. При неполном повреждении постцентральной извилины возникают нарушения кожной чувствительности на противоположной стороне тела, при двустороннем полном повреждении – анестезия (полная потеря чувствительности).
Мышечно-суставная (проприоцептивная) чувствительность проецируется в переднюю (предцентральную) и заднюю (постцентральную) центральные извилины.
Зрительная зона (ядро зрительного анализатора) находится в затылочной доле по краям шпорной борозды. При поражении затылочной доли наступает полная корковая слепота.
Слуховая зона (ядро слухового анализатора) локализуется в верхней височной извилине (поперечные височные извилины, или извилины Р.Гешля) в глубине латеральной борозды. Сюда поступает информация от рецепторов улитки внутреннего уха.
Вкусовая зона расположена в лимбической системе (парагиппокампальной извилине и крючке). Эта область получает импульсацию от вкусовых рецепторов слизистой оболочки полости рта и языка.
Обонятельная зона расположена также в лимбической системе (парагиппокампальной извилине и крючке). Сюда поступают импульсы от обонятельных рецепторов слизистой оболочки полости носа.
Зоны речи.
В коре имеется несколько зон, ведающих функцией речи.
Моторный центр речи (центр П.Брока) находится в лобной доле левого полушария — у «правшей», в лобной доле правого — у «левшей».
Сенсорный центр речи (центр К.Вернике) расположен в височной доле.
10) 3она, обеспечивающая восприятие письменной (зрительной) речи, находится в угловой извилине нижней теменной дольки.
Ассоциативные зоны расположены в теменных, лобных и других долях коры. Они осуществляют связь между различными областями коры, объединяя все поступающие импульсы в целостные акты научения (чтение, речь, письмо), логического мышления, памяти и обеспечивая возможность целесообразной реакции поведения. При нарушении ассоциативных зон появляется агнозия (греч. а — отрицание, gnosis — знание, познание) -неспособность узнавать предметы и апраксия (греч. apraxia — бездействие) — неспособность производить заученные движения.
Долгое время считалось, что левое полушарие (у «правшей») является доминантным (лат. dominans — господствующий), а правое — подчиненным. К настоящему времени имеются данные о функциональной асимметрии полушарий, под которой понимают такое неравенство, при котором в отношении одних функций главным является левое, а в отношении других — правое полушарие. Установлено, что левое полушарие ответственно за речевые функции, логическое и математическое мышление, за формирование положительных эмоций. Правое полушарие отвечает за формирование музыкальных, художественных и других способностей, отрицательных эмоций (печаль, страх и т.д.).
Базальные ядра — это комплекс подкорковых образований: хвостатое ядро, скорлупа, бледный шар, ограда, миндалевидное тело, расположенный в основании больших полушарий вблизи промежуточного мозга и окруженный волокнами внутренней капсулы. Хвостатое ядро и скорлупа чечевицеобразного ядра объединяются под общим названием «полосатое тело» в связи с тем, что скопления нервных клеток, образующих серое вещество, чередуются с прослойками белого вещества. Хвостатое ядро и скорлупа относятся к филогенетически более новым образованиям — неостриатуму (стриатуму). Бледный шар объединяет две светлые мозговые пластинки (латеральную и медиальную) чечевицеобразного ядра, которые иногда называют латеральным и медиальным бледными шарами. Бледный шар является более старым образованием — палеостриатумом (паллидумом). Стриатум (хвостатое ядро и скорлупа) вместе с паллидумом (бледным шаром) образуют стриопаллидарную систему подкорковых ядер.
Для лучшего усвоения и запоминания представим базальные ядра в виде схемы.
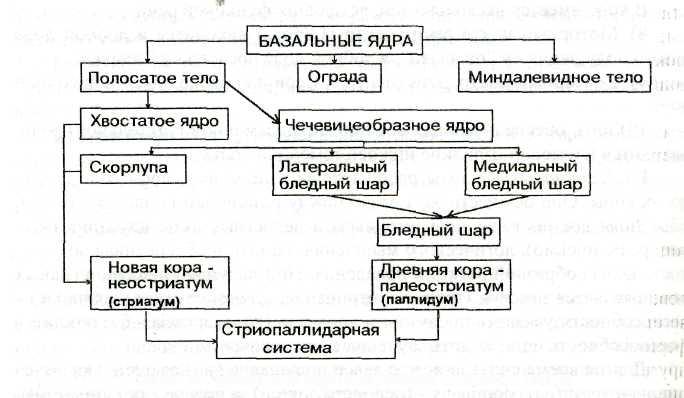
Базальные ядра полушарий большого мозга.
Ядра полосатого тела являются высшими подкорковыми двигательными центрами, входящими в состав экстрапирамидной системы, которая регулирует сложные автоматизированные двигательные акты. К экстрапирамидной системе относятся также черное вещество и красные ядра среднего мозга.
Хвостатое ядро и скорлупа (полосатое тело) регулируют сложные двигательные функции, безусловнорефлекторные реакции цепного характера: бег, плавание, прыжки. Эти функции они осуществляют через бледный шар, притормаживая его деятельность. Кроме того, полосатое тело через гипоталамус регулирует вегетативные функции организма, а также вместе с ядрами промежуточного мозга обеспечивает осуществление безусловных рефлексов — инстинктов.
Бледный шар является центром сложных двигательных рефлекторных реакций (ходьба, бег), формирует сложные мимические реакции, участвует в обеспечении правильного распределения мышечного тонуса. Свои функции бледный шар осуществляет через красные ядра и черное вещество среднего мозга. При раздражении бледного шара наблюдается общее сокращение скелетных мышц на противоположной стороне тела. При поражении бледного шара движения теряют свою плавность, становятся неуклюжими, скованными.
Лимбическая система («висцеральный мозг») — это комплекс образований обонятельного мозга: обонятельная луковица, обонятельный тракт, обонятельный треугольник, переднее продырявленное вещество, расположенный на нижней поверхности лобной доли (периферический отдел обонятельного мозга), а также поясная и парагиппокампальная (вместе с крючком) извилины, зубчатая извилина, гиппокамп (центральный отдел обонятельного мозга) и некоторые другие структуры, расположенные в виде кольца в области нижних отделов коры и окружающие верхнюю часть ствола мозга. Миндалевидное тело, по-видимому, относится к подкорковым обонятельным центрам и к лимбической системе.
Лимбическая система является высшим корковым центром регуляции деятельности вегетативной нервной системы и гипофиза. В ней осуществляется интеграция трех видов информации.»
о деятельности внутренних органов;
обонятельная;
о деятельности чувствительных и двигательных ассоциативных зон коры.
Лимбическая система отвечает за мотивацию и выработку сложных поведенческих актов, успешное выполнение которых требует координации вегетативных и соматических рефлексов. Она активно участвует также в формировании эмоций, памяти, состояний сна, бодрствования и многих других реакций организма. Как филогенетически более древнее образование лимбическая система оказывает регулирующее влияние на кору большого мозга и подкорковые структуры, устанавливая необходимое соответствие уровней их активности. Особенностью лимбической системы является то, что между ее структурами имеются простые двусторонние связи и сложные пути, образующие множество замкнутых кругов. Круги разного функционального назначения связывают лимбическую систему со многими структурами ЦНС, что позволяет последней реализовать функции, специфика которых определяется включенной дополнительной структурой. Так, например, включение хвостатого ядра в один из кругов лимбической системы определяет ее участие в организации тормозных процессов высшей нервной деятельности. Обилие связей лимбической системы со структурами ЦНС затрудняет выделение функций мозга, в которых она не принимала бы участия.
Коре большого мозга свойственна постоянная электрическая активность. Если к поверхности коры или к коже головы приложить два электрода и соединить их с усилителем, то можно записать колебания электрических потенциалов различной формы, амплитуды и частоты. Запись этих колебаний (биопотенциалов) непосредственно от коры называется электрокортикограммой, от кожи головы — электроэнцефалограммой, а сам метод исследования — электроэнцефалографией. Впервые электроэнцефалограмма (ЭЭГ) была зарегистрирована у животных в 1913 году врачом В.В.Правдич-Неминским, у человека — в 1929 году врачом Г.Бергером.
Биоэлектрическую активность головного мозга в функциональном отношении делят на 2 основных вида:
спонтанную (фоновую) активность;
вызванные потенциалы — ответы на фоне спонтанной активности.
Под спонтанной активностью понимают те ритмы, которые регистрируются в покое. Предполагают, что спонтанные волны ЭЭГ являются постсинаптическими потенциалами возбуждения и торможения, преимущественно дендритного (75%) и аксосоматического происхождения (25%). Задающим ритм структурами большинство авторов считает гиппокамп, таламус и ретикулярную формацию, так как стоит только отделить кору от этих образований, как ритм коры исчезает. Физиологический смысл ритма заключается в том, что, если бы нейроны все время работали, они быстро бы истощились.
Различают 4 основных типа ритмов ЭЭГ.
Альфа-ритм — это ритмические колебания потенциалов синусоидальной формы с частотой 8-13 в секунду и амплитудой 20-80 мкВ (микровольт). Регистрируется в условиях покоя при закрытых глазах. Лучше выражен в затылочной области; у слепых людей альфа-ритм может отсутствовать.
Бета-ритм — это потенциалы с частотой колебаний от 14 до 35 в секунду и более низкой амплитудой от 10 до 30 мкВ. Более выражен в лобных долях.
Тета-ритм — потенциалы с частотой колебаний от 4 до 7 в секунду и высокой амплитудой — 100-150 мкВ. Наблюдается во время неглубокого сна, при гипоксии, неглубоком наркозе.
Дельта-ритм — самые медленные волны. Имеет частоту колебаний потенциалов 0,5-3 в секунду, амплитуду 250-300 мкВ (до 1000 мкВ). Наблюдается в состоянии глубокого сна, наркоза, вокруг очага опухоли (локальные дельта-волны с большой амплитудой — дельта-фокус).
Электроэнцефалография широко используется в клинической практике для наблюдения за состоянием головного мозга во время больших oneраций, а также для диагностики ряда заболеваний (эпилепсия, опухоли головного мозга и др.).
11.6.2.3. Строение конечного мозга
Конечный мозг (telencephalon) состоит из двух полушарий большого мозга, разделенных продольной щелью и соединяющихся между собой в глубине этой щели при помощи мозолистого тела, передней и задней спаек, а также спайки свода. Полость конечного мозга образуют правый и левый боковые желудочки, каждый из которых находится в соответствующем полушарии. Полушарие большого мозга состоит из наружных покровов — коры большого мозга (плащ), глубжележащего белого вещества и расположенных в нем скоплений серого вещества — базальных ядер. Граница между конечным и следующим за ним промежуточным мозгом проходит в том месте, где внутренняя капсула прилежит к латеральной стороне таламуса.
Полушарие большого мозга (hemispherium cerehralis) снаружи покрыто тонкой пластинкой серого вещества — корой большого мозга. Каждое полушарие имеет три поверхности: наиболее выпуклую верхнелатеральную; плоскую, обращенную к соседнему полушарию медиальную; нижнюю, которая имеет сложный рельеф, соответствующий внутреннему основанию черепа. Рельеф поверхностей полушарий очень сложен в связи с наличием более или менее глубоких борозд большого мозга и расположенных между ними валикообразных розвышений – извилин. Глубина, протяженность боррзд и выпуклых извилин, их форма и направление очень изменчивы.
Верхнелатеральная поверхность полушария. В переднем отделе каждого полушария большого мозга находится лобная доля, ограниченная снизу латеральной бороздой (сильвиева борозда), а сзади — глубокой центральной бороздой (рис. 11.25). Спереди от центральной борозды, почти параллельно ей, располагается предцентральная борозда, от которой вперед направляются верхняя и нижняя лобные борозды, которые делят верхнелатеральную поверхность лобной доли на извилины. Между центральной бороздой сзади и предцентральной бороздой спереди находится предцентральная извилина. Выше верхней лобной борозды лежит верхняя лобная извилина. Между верхней и нижней лобными бороздами тянется средняя лобная извилина
Покрьшечная часть (лобная покрьшка) расположена между восходящей ветвью и нижним отделом предцентральной борозды. Эта часть лобной доли получила такое название потому, что она прикрывает лежащую в глубине борозды островковую долю (островок).
Кзади от центральной борозды находится теменная доля. Задней границей этой доли является теменно-затылочкая борозда. Нижней границей теменной доли является латеральная борозда (ее задняя ветвь), отделяющая эту долю (ее передние отделы) от височной. В пределах теменной доли выделяют постцентралькую борозду, которая лежит позади центральной борозды и почти параллельно ей. Между центральной и постцентральной бороздами располагается постцентральная извилина. Вверху она переходит на медиальную поверхность полушария большого мозга, где соединяется с предцентральной извилиной лобной доли, образуя вместе с нею парацентральную дольку. От постцентральной борозды кзади отходит внутритеменная борозда. Она параллельна верхнему краю полушария. Кверху от внутритеменной борозды находится группа мелких извилин, получивших название верхней теменной дольки.
Рис. 11.25. Латеральная поверхность полушарий.
Ниже этой борозды лежит нижняя теменная долька, в пределах которой выделяют две извилины: надкраевую и угловую. Нижняя часть нижней теменной дольки и прилежащие к ней нижние отделы постцентральной извилины вместе с нижней частью предцентральной извилины, нависающие над остров-ковой долей, образуют лобно-теменную покрышку островка.
Затылочная доля располагается позади теменно-затылочной борозды и ее условного продолжения на верхнелатеральной поверхности полушария. Затылочная доля кончается затылочным полюсом. Борозды и извилины на верхнелатеральной поверхности затылочной доли очень вариабельны. Наиболее часто и лучше других выражена поперечная затылочная борозда.
Височная доля занимает нижнебоковые отделы полушария и отделяется от лобной и теменной долей глубокой латеральной бороздой. Край височной доли, прикрывающий островковую долю, получил название височной покрышки. На боковой поверхности височной доли видны две борозды — верхняя и нижняя височные. Извилины височной доли ориентированы вдоль борозд. Верхняя височная извилина расположена между латеральной бороздой вверху и верхней височной внизу. На верхней поверхности этой извилины, скрытой в глубине латеральной борозды, располагаются две-три короткие поперечные височные извилины (извилины Гешля), разделенные поперечными височными бороздами. Между верхней и нижней височными бороздами находится средняя височная извилина. Нижнелатеральный край височной доли занимает нижняя височная извилина, ограниченная сверху одноименной бороздой. Задний конец этой извилины продолжается в затылочную долю.
Островковая доля (островок, insula), находится в глубине латеральной борозды. Глубокая круговая борозда островка отделяет островок от окружающих его отделов мозга. На поверхности островка имеются извилины островка, длинная и короткие
Медиальная поверхность полушария. Все доли полушария, за исключением островковой, принимают участие в образовании его медиальной поверхности. Над мозолистым телом находится борозда мозолистого тела, которая продолжается в борозду гиппокампа.
Выше борозды мозолистого тела находится поясная борозда. На уровне валика мозолистого тела от поясной борозды вверх ответвляется краевая часть. Между бороздой мозолистого тела и поясной бороздой находится поясная извилина, охватывающая мозолистое тело спереди, сверху и сзади. Сзади и книзу от валика мозолистого тела поясная извилина суживается, образуя перешеек поясной извилины, которая книзу переходит в более широкую извилину гиппокампа. Поясная извилина, перешеек и парагиппокампальная извилина известны как сводчатая извилина. В глубине борозды гиппокампа находится довольно тонкая полоска серого цвета, разделенная мелкими поперечными бороздками – зубчатая извилина. Участок медиальной поверхности полушария, находящийся между поясной бороздой и верхним краем полушария, относится к лобной и теменной долям.
Кпереди от верхнего края центральной борозды находится медиальная поверхность верхней лобной извилины, а непосредственно к указанному участку центральной борозды прилежит парацентральная долька. Между краевой частью спереди и тененно-затылочной бороздой сзади находится предклинье, принадлежащий теменной доле участок полушария.
На медиальной поверхности затылочной доли расположены сливающиеся друг с другом под острым углом, открытым кзади, две глубокие борозды: теменно-затылочная борозда, отделяющая теменную долю от затылочной, и шпорная борозда. Участок затылочной доли, лежащий между теменно-затылочной и шпорной бороздами называется клином, Шпорная борозда ограничивает сверху язычную извилину, снизу от которой располагается коллатеральная борозда.
Нижняя поверхность полушария. Передние отделы этой поверхности образованы лобной долей полушария, позади которой выступает височный полюс, а также находятся нижние поверхности височной и затылочной долей, переходящие одна в другую без заметных границ. На нижней поверхности лобной доли, несколько латеральнее и параллельно продольной щели большого мозга, находится обонятельная борозда. Снизу к ней прилежат обонятельная луковица и обонятельный тракт, переходящий сзади в обонятельный треугольник, в области которого видны медиальная и латеральная обонятельные полоски. Участок лобной доли между продольной щелью большого мозга и обонятельной бороздой получил название прямой извилины. Поверхность лобной доли, лежащая латеральнее обонятельной борозды, разделена неглубокими глазничными бороздами.
В заднем отделе нижней поверхности полушария хорошо различима коллатеральная борозда, лежащая книзу и латерально от язычной извилины на нижней поверхности затылочной и височной долей, латерально от парагиппокампальной извилины. Несколько кпереди от переднего конца коллатеральной борозды находится носовая борозда. Латеральнее коллатеральной борозды лежит медиальная затылочно-височная извилина. Между этой извилиной и расположенной кнаружи от нее латеральной затылочно-височной извилиной находится затылочно-височная борозда,
Ряд отделов головного мозга, расположенных преимущественно на медиальной поверхности полушария и являющихся субстратом для формирования таких общих состояний, как бодрствование, сон, эмоции, мотивации поведения и др., выделяют под названием «лимбическая система» (рис. 11.26).
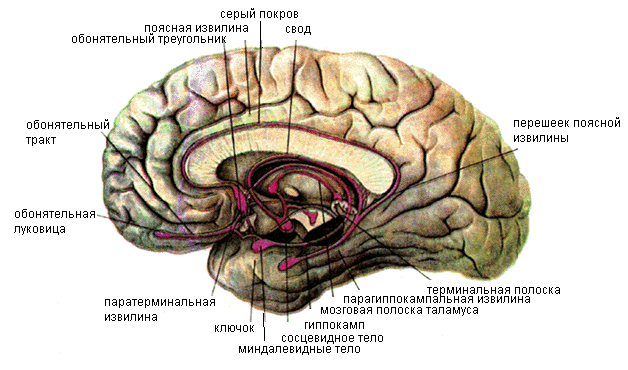
Рис. 11.26. Структуры лимбической системы головного мозга.
Поскольку эти реакции сформировались в связи с первичными функциями обоняния (в филогенезе), их морфологической основой являются отделы мозга, которые развиваются из нижнєлатеральных отделов мозгового пузыря и относятся к так называемому обонятельному мозгу (rhinencephalon). Лимбическую систему составляют обонятельная луковица, обонятельный тракт, обонятельный треугольник, переднее продырявленное вещество, расположенные на нижней поверхности лобной доли (периферический отдел обонятельного мозга), а также поясная и парагиппокампальная (вместе с крючком) извилины, зубчатая извилина, гиппокамп (центральный отдел обонятельного мозга) и некоторые другие структуры.
Кора большого мозга (плащ) (cortex cerebri, pallium) представлена серым веществом, расположенным по периферии полушарий большого мозга. Плошаль поверхности коры одного полушария у взрослого человека в среднем равна 220 000 мм2, причем на видимые части извилин приходится 1/3, а на боковые и нижние стенки борозд – 2/3 всей площади коры. Толщина коры в различных участках неодинакова и колеблется от 1,5 до 5,0 мм. Наибольшая толщина отмечается в верхних участках предцентральной и постцентральной извилин и парацентральной дольки.
Как показал В.А. Бец, не только вид нервных клеток, но и их взаиморасположение неодинаково в различных участках коры. Волоконное строение коры (миелоархитектоника) в основном соответствует клеточному ее составу (цитоархитектоника). Типичным для новой (neocortex) коры большого мозга взрослого человека является расположение нервных клеток в виде шести слоев (пластинок). На медиальной и нижней поверхностях полушарий большого мозга сохранились участки старой (archicortex) и древней (paleocortex) коры, имеющей двухслойное и трехслойное строение. 1) молекулярная пластинка, наружная зернистая пластинка, 3) наружная пирамидная пластинка (слой малых, средних пирамид), внутренняя зернистая пластинка, внутренняя пирамидная пластинка (слой больших пирамид, или клеток Беца), мультиформная (полиформная) пластинка (рис. 11.27).
Исследования, проведенные учеными разных стран в конце XIX – начале XX в., позволили создать цитоархитектонические карты коры большого мозга человека и животных, в основу которых были положены особенности строения коры в каждом участке полушария. К. Бродман выделил в коре 52 цитоархитектонических поля, Ф. Фогт и О. Фогт с учетом волоконного строения описали в коре большого мозга 150 миелоархитектонических участков.
Локализация функций в коре полушарий большого мозга
В коре большого мозга располагаются центры, регулирующие выполнение тех или иных функций. Морфологическим подтверждением данных физиологии и клиники явилось учение о разнокачественности строения коры полушарий большого мозга в различных ее участках — цито- и миелоархитектоника коры. И.П. Павлов рассматривал кору полушарий большого мозга как сплошную воспринимающую поверхность, как совокупность корковых концов анализаторов. Различные анализаторы тесно взаимосвязаны, в связи с этим в коре большого мозга осуществляются анализ и синтез, выработка ответных реакций, регулирующих любые виды деятельности человека.
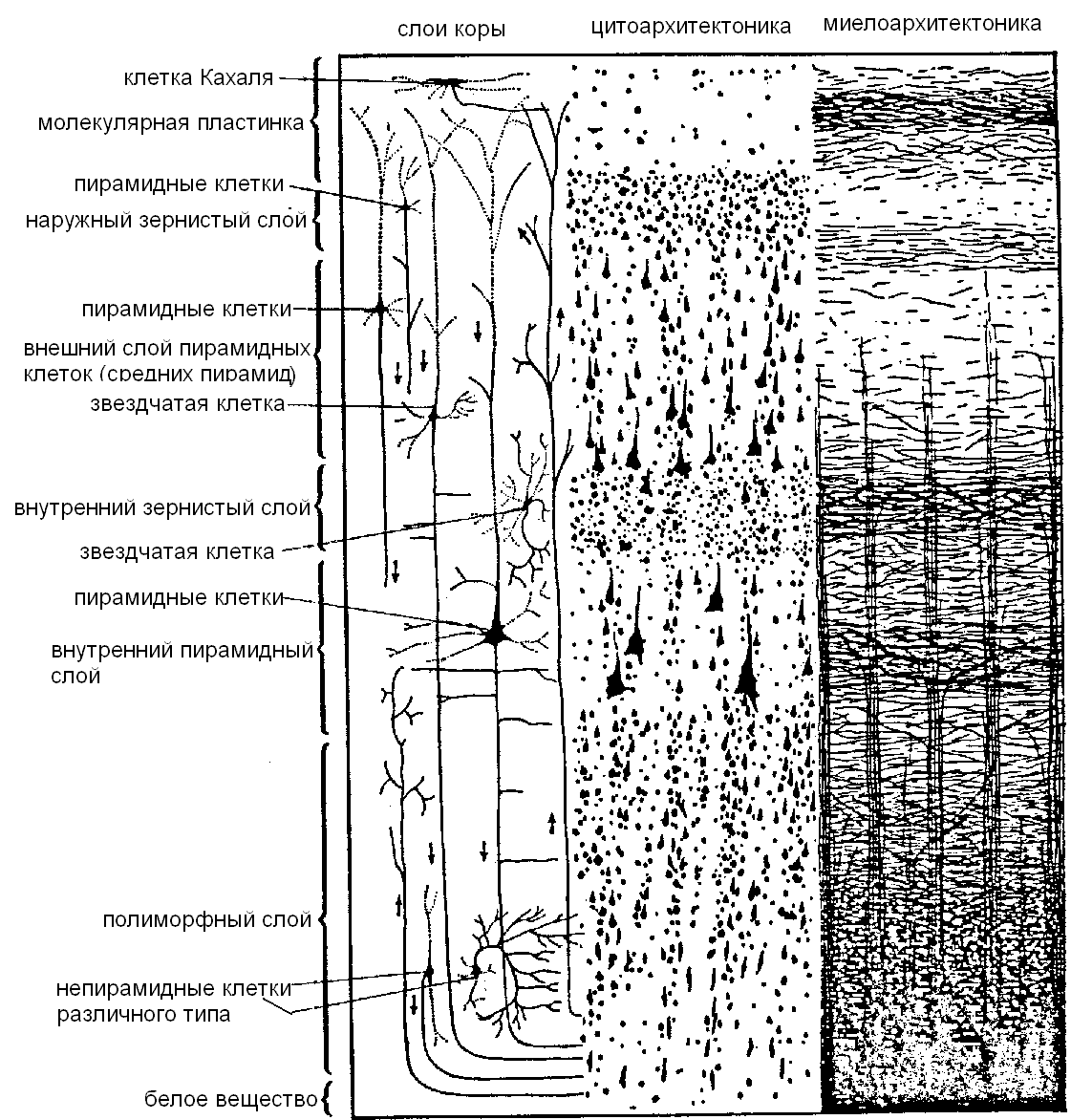
Рис. 11.27. Строение коры больших полушарий.
В коре большого мозга различают ядро и рассеянные элементы. Ядро — это место концентрации нервных клеток коры, составляющих точную проекцию всех элементов определенного периферического рецептора, где происходят высший анализ, синтез и интеграция функций. Рассеянные элементы могут располагаться как по периферии ядра, так и на значительном расстоянии от него. В них совершаются более простые анализ и синтез. Наличие рассеянных элементов при разрушении (повреждении) ядра отчасти позволяет компенсировать нарушенную функцию. Площади, занимаемые рассеянными элементами различных анализаторов, могут наслаиваться друг на друга, перекрывать друг друга.
Рассмотрим положение некоторых корковых концов различных анализаторов (ядер) по отношению к извилинам и долям полушарий большого мозга у человека (рис. 11.28).
В коре постцентральной извилины (поля 1, 2, 3) и верхней теменной дольки (поля 5 и 7) залегают нервные клетки, образующие ядро коркового анализатора общей чувствительности (температурной, болевой, осязательной) и проприоцептивной. Проводящие чувствительные пути, перекрещиваются либо на уровне различных сегментов спинного мозга (пути болевой, температурной чувствительности, осязания и давления), либо на уровне продолговатого мозга (пути проприоцептивной чувствительности коркового направления). Вследствие этого постцентральные извилины каждого из полушарий связаны с противоположной половиной тела. В постцентральной извилине все рецепторные поля различных участков тела человека спроецированы таким образом, что наиболее высоко расположены корковые концы анализатора чувствительности нижних отделов туловища и нижних конечностей, а наиболее низко (ближе к латеральной борозде) проецируются рецепторные поля верхних участков тела и головы, верхних конечностей.
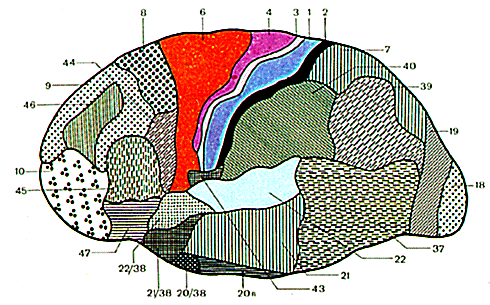
Рис. 11.28. Цитоархитектонические поля левого полушария большого мозга (верхелатеральная поверхность).
Ядро двигательного анализатора находится в основном в двигательной области коры, к которой относятся предцентральная извилина (поля 4 и 6) и парацентральная долька на медиальной поверхности полушария. Причем в верхних участках предцентральной извилины и в парацентральной дольке расположены клетки, импульсы от которых направляются к мышцам самых нижних отделов туловища и нижних конечностей. В нижней части предцентральной извилины находятся двигательные центры, регулирующие деятельность мышц лица.
Ядро анализатора, обеспечивающее функцию сочетанного поворота головы и глаз в противоположную сторону, находится в задних отделах средней лобной извилины, в премоторной зоне (поле 8). Сочетанный поворот глаз и головы регулируется не только при поступлении в кору лобной извилины проприоцептивных импульсов от мышц глазного яблока, но и при поступлении импульсов с сетчатки глаза в поле 17, находящееся в затылочной доле, по соседству с ядром зрительного анализатора.
В надкраевой извилине находится ядро двигательного анализатора, функциональное значение которого состоит в осуществлении синтеза всех целенаправленных сложных комбинированных движений. Это ядро асимметрично. У правшей оно находится в левом, а у левшей — в правом полушарии. Осуществление целенаправленных движений происходит за счет образования временных связей между клетками, расположенными в предцентральной и надкраевой извилинах. Поражение поля 40 не вызывает паралича, а лишь приводит к потере способности производить сложные координированные целенаправленные движения — апраксии.
В коре верхней теменной дольки (поле 7) находится ядро кожного анализатора, которому присуща функция узнавания предметов на ощупь (стреогнозии). Корковый конец этого анализатора находится в правом полушарии и представляет собой проекцию рецепторных полей левой верхней конечности. Для правой верхней конечности ядро анализатора находится в левом полушарии.
В глубине латеральной борозды, находится ядро слухового анализатора (поля 41, 42, 52). К нервным клеткам, составляющим ядро слухового анализатора каждого из полушарий, проходят проводящие пути от рецепторов как левой, так и правой стороны. В связи с этим одностороннее поражение этого ядра не вызывает полной утраты способности воспринимать звуки. Двустороннее поражение сопровождается «корковой глухотой».
Ядро зрительного анализатора располагается на медиальной поверхности затылочной доли полушария большого мозга, по обеим сторонам от шпорной борозды (поля 17, 18, 19). Ядро зрительного анализатора правого полушария связано проводящими путями с латеральной половиной сетчатки правого глаза и медиальной половиной сетчатки левого глаза. В коре затылочной доли левого полушария проецируютея соответственно рецепторы латеральной половины сетчатки левого глаза и медиальной половины сетчатки правого глаза. Как и для ядра слухового анализатора, только двустороннее поражение ядер зрительного анализатора приводит к полной «корковой слепоте». Поражение поля 18, находящегося несколько выше поля 17, сопровождается потерей зрительной памяти, однако утрата зрения не отмечается. Наиболее высоко по отношению к двум предыдущим в коре затылочной доли находится поле 19, поражение которого сопровождается утратой способности ориентироваться в незнакомой окружающей обстановке.
На нижней поверхности височной доли в области крючка и отчасти в области гиппокампа (поле II), находится ядро обонятельного анализатора. Эти участки с точки зрения филогенеза относятся к наиболее древним частям коры большого мозга. Чувство обоняния и чувство вкуса тесно взаимосвязаны, что объясняется близким расположением ядер обонятельного и вкусового анализаторов. Вкусовое восприятие нарушается при поражении коры самых нижних отделов постцентральной извилины (поле 43)..
Описанные корковые концы некоторых анализаторов имеются в коре полушарий большого мозга не только человека, но и животных. Они специализированы на восприятии, анализе и синтезе сигналов, поступающих из внешней и внутренней среды, составляющих, по определению И.П. Павлова, первую сигнальную систему действительности.
Вторая сигнальная система имеется только у человека и обусловлена развитием речи.
Анализаторы зрительного и слухового восприятия речевых сигналов расположены по соседству с анализаторами зрения и слуха. Следует указать на то обстоятельство, что речевые анализаторы у правшей локализуются в левом полушарии, а у левшей — в правом. Рассмотрим положение в коре большого мозга некоторых из речевых анализаторов.
Ядро двигательного анализатора письменной речи находится в заднем отделе средней лобной извилины (поле 40). Разрушение поля 40 не приводит к нарушению всех видов движений, а сопровождается лишь утратой способности производить рукой точные и тонкие движения при начертании букв, знаков и слов (аграфия).
Ядро двигательного анализатора артикуляции речи (речедвигательный анализатор) располагается в задних отделах нижней лобной извилины (поле 44, или центр Брока). Повреждение этой области приводит к двигательной афазии, т.е. утрате способности произносить слова. При этом не утрачивается способность к произношению звуков или пению.
В центральных отделах нижней лобной извилины (поле 45) находится ядро речевого анализатора, связанного с пением. Поражение поля 45 сопровождается вокальной амузией- неспособностью к составлению и воспроизведению музыкальных фраз и аграмиатизном, когда утрачивается способность к составлению осмысленных предложений из отдельных слов. Речь таких больных состоит из несвязанного по смысловому значению набора слов.
Ядро слухового анализатора устной речи располагается в области верхней височной извилины. (поле 42). Поражение ядра не нарушает слухового восприятия звуков вообще, однако утрачивается способность понимать слова, речь (словесная глухота, или сенсорная афазия).
В средней трети верхней височной извилины (поле 22) находится ядро коркового анализатора, поражение которого сопровождается наступлением музыкальной глухоты: музыкальные фразы воспринимаются как бессмысленный набор различных шумов.
Ядро зрительного анализатора письменной речи (поле 39) расположено в угловой извилине нижней теменной дольки. Поражение этого ядра приводит к утрате способности воспринимать написанный текст, читать (алексия).
Базальные (подкорковые) ядра и белое вещество конечного мозга
Помимо коры серое вещество в каждом из полушарий большого мозга залегает в виде отдельных ядер, или узлов. Эти узлы находятся в толще белого вещества, ближе к основанию мозга. Скопления серого вещества в связи с их положением получили наименование базальных (подкорковых, центральных) ядер (узлов) (nuclei basales). К базальным ядрам полушарий относят полосатое тело, состоящее из хвостатого и чечевицеобразного ядер; ограду и миндалевидное тело (рис. 11.29, 11.30).
Полосатое тело (corpus striatum) на горизонтальных и фронтальных разрезах мозга имеет вид чередующихся полос серого и белого вещества. Наиболее медиально и впереди находится хвостатое ядро (nucleus caudatus), от которого отделяет полоска белого вещества – колено внутренней капсулы. Передний отдел хвостатого ядра утолщен и образует головку, которая составляет латеральную стенку переднего рога бокового желудочка. Располагаясь в лобной доле полушария, головка хвостатого ядра внизу примыкает к переднему продырявленному веществу. В этом месте головка хвостатого ядра соединяется с чечевицеобразным ядром. Суживаясь кзади, головка продолжается в более тонкое тело. Задний отдел хвостатого ядра — квост постепенно истончается, изгибается книзу, участвует в образовании верхней стенки нижнего рога бокового желудочка и достигает миндалевидного тела, лежащего в переднемедиальных отделах височной доли (кзади от переднего продырявленного вещества). Латеральнее головки хвостатого ядра находится прослойка белого вещества — передняя ножка (бедро) внутренней капсулы, отделяющая это ядро от чечевицеобразного.
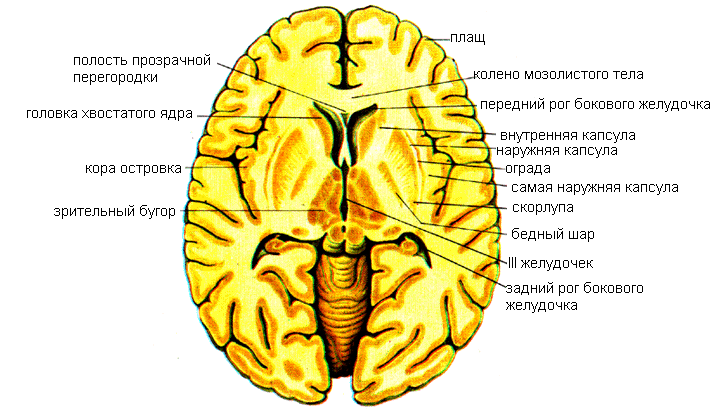
Рис. 11.29. Горизонтальный разрез головного мозга. Базальные ганглии.
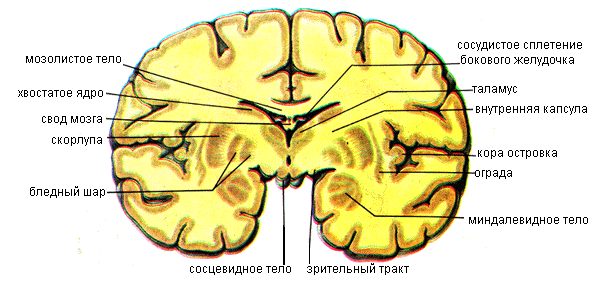
Рис. 11.30. Фронтальный разрез головного мозга на уровне сосцевидных тел.
Чечевицеобразное ядро (nucleus lentiformis) находится латеральнее таламуса и хвостатого ядра. Нижняя поверхность переднего отдела чечевицеобразного ядра прилежит к переднему продырявленному веществу и соединяется с хвостатым ядром. Медиальная часть чечевицеобразного ядра на горизонтальном разрезе головного мозга суживается и углом обращена к колену внутренней капсулы, находящемуся на границе таламуса и головки хвостатого ядра.
Две параллельные вертикальные прослойки белого вещества, расположенные почти в сагиттальной плоскости, делят чечевицеобразное ядро на три части. Наиболее латерально лежит скорлупа, имеющая более темную окраску. Медиальнее скорлупы расположены две светлые мозговые пластинки – медиальная и латеральная – бледный шар.
Хвостатое ядро и скорлупа относятся к филогенетически более новым образованиям (neostriatum). Бледный шар является более старым образованием paleostriatum.
Ограда (claustrum) расположена в белом веществе полушария, сбоку от скорлупы, между последней и корой островковой доли, имеет вид тонкой вертикальной пластинки серого вещества. От скорлупы ее отделяет прослойка белого вещества –наружная капсула.
Миндалевидное тело (corpus amygdaloideum) находится в белом веществе височной доли полушария.
Белое вещество полушарий большого мозга представлено различными системами нервных волокон, среди которых выделяют ассоциативные, комиссуральные, проекционные.
Ассоциативные нервные волокна, которые выходят из коры полушария (экстракортикальные), располагаются в пределах одного полушария, соединяя различные функциональные центры. Комиссуральные нервные волокна проходят через спайки мозга (мозолистое тело, передняя спайка). Проекционные нервные волокна, идущие от полушария большого мозга к нижележащим его отделам и к спинному мозгу, а также следующие в обратном направлении от этих образований, составляют внутреннюю капсулу и ее лучистый венец.
Внутренняя капсула (capsula interna) — это толстая изогнутая под углом пластинка белого вещества. С латеральной стороны она ограничена чечевицеобразным ядром, а с медиальной — головкой хвостатого ядра (спереди) и таламусом (сзади). Внутреннюю капсулу подразделяют на три отдела. Между хвостатым и чечевицеобразным ядрами находится передняя ножка внутренней капсулы; между таламусом и чечевицеобразным ядром — задняя ножка внутренней капсулы. Место соединения этих двух отделов под углом, открытым латерально, составляет колено внутренней капсулы. В колене внутренней капсулы располагаются волокна нисходящих и восходящих путей.
Мозолистое тело (corpus callosum) содержит комиссуральные проводящие пути, переходящие из одного полушария в другое и соединяющие участки коры, принадлежащие правому и левому полушариям, с целью объединения На сагиттальном разрезе головного мозга можно различить изгибы и части мозолистого тела: колено, продолжающееся книзу в клюв, а затем в терминальную (концевую) пластинку. Среднюю часть называют стволом мозолистого тела. Кзади ствол продолжается в утолщенную часть – валик. Поперечно идущие волокна мозолистого тела в каждом полушарии большого мозга образуют лучистость мозолистого тела.
Под мозолистым телом находится свод (forni), который состоит из двух дугообразно изогнутых тяжей, соединенных в средней своей части при помощи поперечно идущих волокон – спайки. Средняя часть свода носит название тела, кпереди и книзу оно продолжается в округлый парный тяж – столб свода. Кзади тело свода продолжается в также парный плоский тяж — ножку свода, сращенную с нижней поверхностью мозолистого тела.
Впереди свода в сагиттальной плоскости располагается прозрачная перегородка (septum pellucidum), которая состоит из двух пластинок, лежащих параллельно друг другу. Между пластинками прозрачной перегородки находится щелевидная полость, содержащая, прозрачную жидкость. Впереди столбов свода находится передняя
Боковой желудочек (ventriculus lateralis) расположен в толще полушария. Различают два боковых желудочка: левый (первый), и правый (второй). Полость желудочка имеет сложную форму (рис. 11.31). Такая форма обусловлена тем, что отделы желудочка располагаются во всех долях полушария (за исключением островка). Теменной доле полушария большого мозга соответствует центральная часть бокового желудочка, лобной доле — передний (лобный) рог, затылочной — задний (затылочный) рог, височной доле — нижний (височный) рог. Центральная часть при помощи межжелудкового отверстия соединяется с III желудочком.
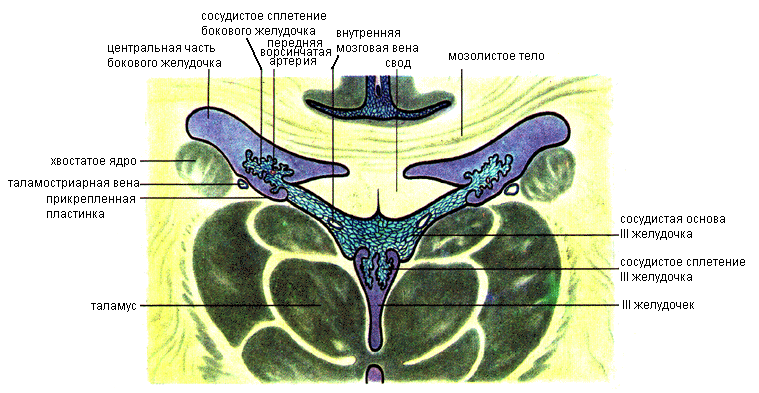
Рис. 11.31. Фронтальный разрез головного мозга на уровне центральной части боковых желудочков.
Оболочки головного мозга
Головной мозг, как и спинной, окружен тремя соединительноткаными листками, или оболочками, являющимися продолжением оболочек спинного мозга, каждая из которых отделена от соседних межоболочечным пространством (рис. 11.32).
Твердая оболочка головного мозга, образованная волокнистой соединительной тканью отличается по строению от аналогичной оболочки спинного мозга. Она является одновременно надкостницей на внутренней поверхности костей черепа, с которыми связана непрочно. В области основания черепа оболочка дает ряд отростков, проникающих в щели и отверстия костей черепа, чем объясняется большая прочность прикрепления здесь твердой оболочки головного мозга. Более того, в местах выхода из полости черепных нервов твердая оболочка головного мозга на некотором протяжении продолжает окружать нерв, образуя его влагалище и проникая вместе с нервом через отверстие наружу.
На внутренней поверхности твердой оболочки различают несколько отростков, которые проникают в продольную щель большого мозга и отделяют друг от друга его полушария. Один из них — серп большого мозга — расположен между двумя полушариями (рис. 11.33). Другой — палатка мозжечка — проходит горизонтально, отделяя мозжечок от большого мозга. Еще одна складка, диафрагма седла, изолирует и защищает гипофиз от давлений всей вышележащей массы мозга.
В определенных участках твердой оболочки головного мозга имеются расщепления, выстланные изнутри эндотелием, — это синусы твердой оболочки головного мозга, по которым оттекает венозная кровь. Особенностью синусов является прочность стенок, что объясняет невозможность их спадения. Кроме того, синусы соединяются с наружными венами головы через эмиссарные вены.
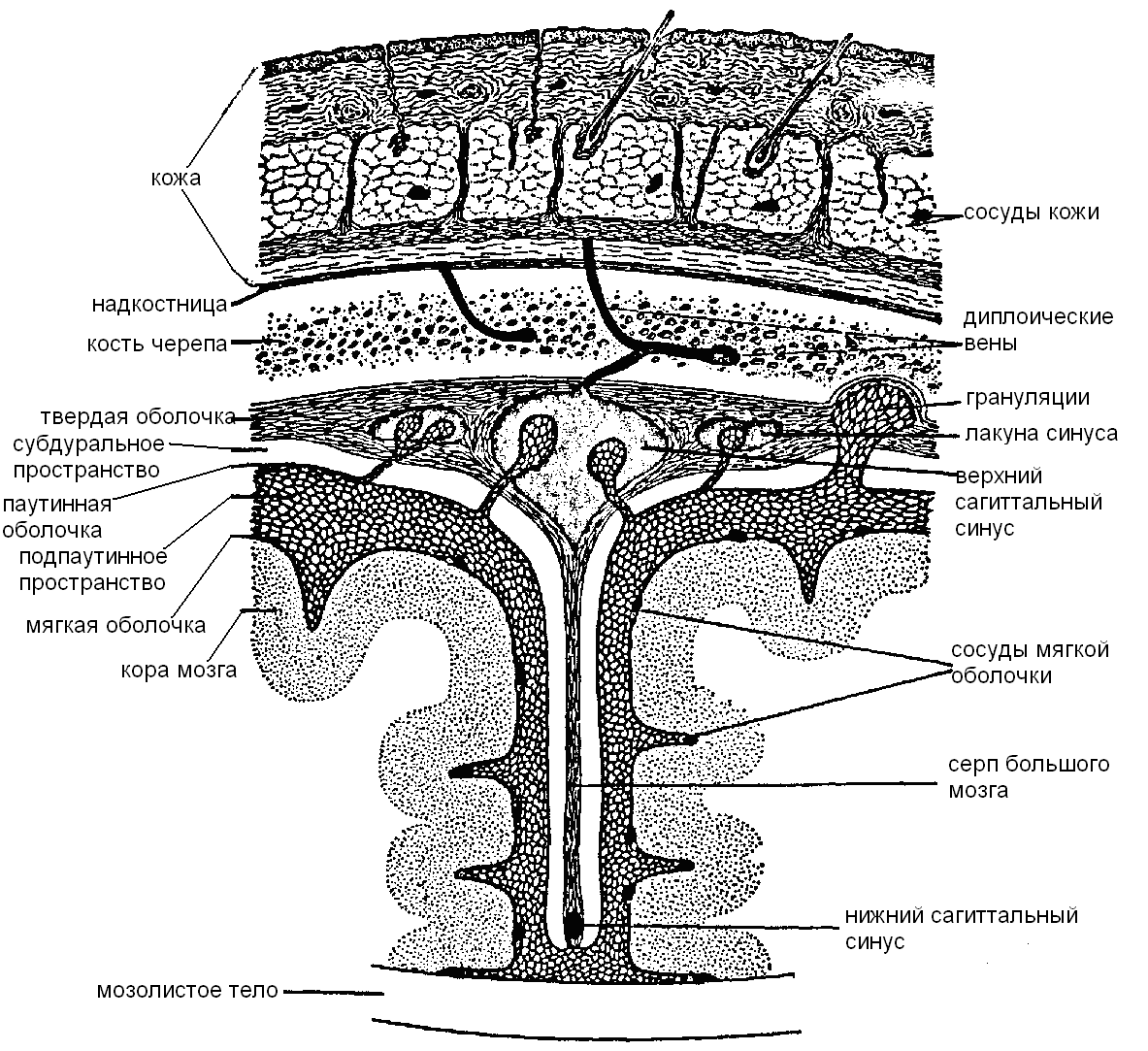
Рис. 11.32. Оболочки головного мозга на фронтальном разрезе.
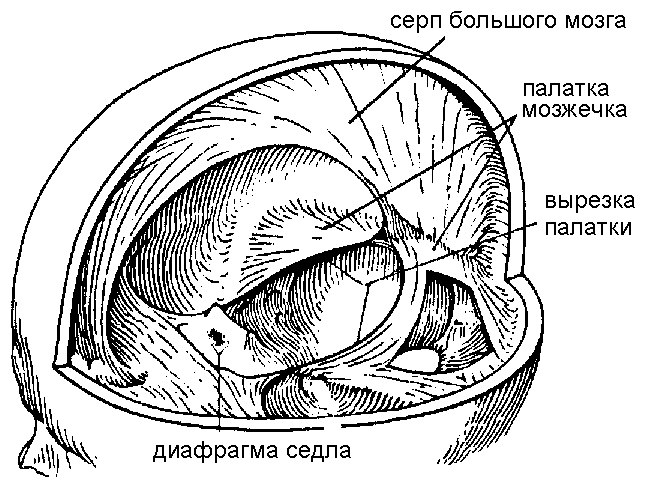
Рис. 11.33. Серп мозга и палатка.
Паутинная оболочкаголовного мозга, образованная эластичной волокнистой тканью, располагается внутри от твердой мозговой и отделена от нее субдуральным пространством. Она отделена от твердой промежутком – субдуральным пространством, которое в норме не существует, так как паутинная оболочка плотно прилежит к твердой (“потенциальное пространство”). Паутинная оболочка соединена с лежащей глубже мягкой оболочкой посредством тонких перемычек. Эти две оболочки отделены друг от друга подпаутинным пространством, заполненным спинномозговой жидкостью. В некоторых отделах подпаутинное пространство расширяется, образуя цистерны. Подпаутинное пространство головного мозга в области большого затылочного отверстия сообщается с подпаутинным пространством спинного мозга.
Цистерны находятся над каждой бороздой или щелью поверхности мозга. Самая крупная из них — большая цистерна— находится между задне-нижней поверхностью мозжечка и продолговатым мозгом. Через твердую мозговую оболочку в интрадуральные венозные синусы проникают небольшие особые выросты паутинной оболочки, покрытые “шапочкой” из эпителиальных клеток. Этогрануляциипаутинной оболочки, через которые часть спинномозговой жидкости, заполняющей субарахноидальное пространство, может поступать в кровяное русло. Эти выросты вдаются в синусы твердой оболочки. На внутренней поверхности костей черепа в месте расположения грануляций отмечаются вдавления и ямочки.
Общепризнанным является мнение об участии грануляции паутинной оболочки в обеспечении оттока спинномозговой жидкости в венозное русло.
Мягкая (сосудистая) оболочка — это самая внутренняя из оболочек головного мозга. Она состоит из соединительной ткани, образующей два слоя (внутренний и наружный), между которыми залегают кровеносные сосуды. Оболочка сращена с наружной поверхность мозга и глубоко проникает во все его щели и борозды. Кровеносные сосуды , покидая сосудистую оболочку, направляются в ткань мозга, обеспечивая его питание. В определенных местах сосудистая оболочка проникает в полости желудочков мозга и образует сосудистые сплетения, проецирующие спинномозговую жидкость.
Система желудочков мозга
Боковые, латеральные желудочки мозга (правый и левый) лежат внутри обоих полушарий большого мозга. В каждом из боковых желудочков различают части: передний, задний и нижний рога, центральная часть. Каждая из них соответствует одной из долей полушария большого мозга: передний рог залегает в лобной доле, задний — в затылочной, нижний — в височной, центральная часть — в теменной. Третий желудочек мозга располагается в срединной сагиттальной плоскости и сообщается с боковыми желудочками посредством впереди лежащих межжелудочковых отверстий, с четвертым желудочком — посредством водопровода мозга. Боковые стенки третьего желудочка образованы медиальными поверхностями зрительных бугров. Четвертый желудочек внизу сообщается с полостью спинного мозга его спинномозговым каналом. В задних отделах четвертого желудочка находятся два отверстия, через которые полость желудочка сообщается с субарахноидальным пространством, — боковые отверстия четвертого желудочка (отверстия Люшка). В самой каудальной части четвертого желудочка есть участок, где его крыша сильно истончается и в ней образуется отверстие, через которое может выходить часть спинномозговой жидкости (срединное отверстие четвертого желудочка, или отверстие Мажанди).
В желудочках мозга находится сосудистое сплетение, выполняющее функцию секреции цереброспинальной жидкости. Большая его часть равномерно распределена между четвертым и латеральными желудочками. Около 10% сосудистого сплетения расположено в третьем желудочке. Во взрослом мозге на долю сосудистого сплетения приходится, как правило, не более 0,25% массы всего мозга (у человека 2 — 3 г).
Ликвор. Головной мозг содержит в среднем 130–140 мл ликвора. Из этого количества жидкости около 30 мл содержится в желудочках мозга, а остальная часть заполняет цистернальные и субарахно-идальные пространства головного и спинного мозга. Эти полости соединяются друг с другом, и давление в них является одной из важных констант организма.
Ликвор – бесцветная жидкость с плотностью 1,006–1,007, со слабощелочной реакцией, рН 7,6. В нем содержится большое количество электролитов и незначительное количество белков, гормонов и ферментов.
Состав церебро-спинальной жидкости в разных отделах головного и спинного мозга несколько отличается по цитологическим показателям и плотности.
Ликвор образуется несколькими путями. В секреции ликвора принимают участие сосудистые сплетения желудочков мозга, сосуды менингеальной оболочки и эпендимы желудочков, а также клетки паренхимы мозга. Секреция и выделение ликвора, а также обратное всасывание происходят постоянно.
Основная масса спинномозговой жидкости образуется в латеральных желудочках мозга, а небольшие количества – в третьем и четвертом желудочках. Основной ток ликвора идет в каудальном направлении, а из желудочков мозга спинномозговая жидкость поступает в субарахноидальное пространство и центральный канал спинного мозга через отверстие Люшка и Мажанди. Ликвор, находящийся в различных отделах головного и спинного мозга (желудочках, субарахноидальном и периваскулярном пространствах), подвергается достоянному перераспределению и колебательному перемещению, которые взаимосвязаны с дыхательными, мышечными и пульсовыми движениями.
Функции ликвора многообразны. Известно, что функциональное состояние и деятельность головного мозга в значительной мере зависят от состава, физических и биологических свойств спинномозговой жидкости. Защитная функция ликвора проявляется в удалении продуктов распада нервных клеток и токсических веществ, идущих по системе нейрон – глия – эпендима – жел. мозг. Поддерживая постоянство состава среды, в которой функционируют нейроны мозга, церебро-спинальная жидкость выполняет гомеостатическую функцию. До настоящего времени не утратила своего значения теория, доказывающая значение ликвора как амортизирующего фактора, выполняющего роль «водяной подушки», обеспечивающей защиту нервных клеток от механических воздействий. Головной и спинной мозг как бы «плавают» в ликворе, и любое механическое воздействие, в частности удары умеренной силы, не вызывают повреждения вещества мозга.
доли, борозды, извилины Серое и белое вещество. Полость полушарий.
Головной мозг состоит из переднего мозга (конечного и промежуточного), среднего мозга, ромбовидного мозга (задний мозг – мост, мозжечок, продолговатый мозг). Между ромбовидным и средним мозгом расположен перешеек ромбовидного мозга.
Головной мозг находится в полости мозгового черепа. Имеет выпуклую верхнелатеральную поверхность и нижнюю поверхность и уплощенную – основание головного мозга.
Масса мозга взрослого человека составляет от 1100 до 2000 грамм; от 20 до 60 лет масса m и объем V остаются максимальными и постоянными, после 60 лет несколько уменьшается. Ни абсолютная, ни относительная масса мозга не является показателем степени умственного развития. Масса мозга Тургенева была 2012 г, Байрона – 2238 г, Кювье – 1830 г, Шиллера – 1871 г, Менделеева – 1579 г, Павлова – 1653 г. Головной мозг состоит из тел нейронов, нервных трактов и кровеносных сосудов. Головной мозг состоит из 3 частей: полушарий большого мозга, мозжечка и мозгового ствола.
Полушария большого мозга достигают максимального развития у человека, позднее других отделов.
Большой мозг состоит из двух полушарий – правого и левого, которые связаны одно с другим толстой спайкой (комиссурой) – мозолистым телом. Правое и левое полушария делятся с помощью продольной щели. Под комиссурой находится свод, представляющий собой два изогнутых волокнистых тяжа, которые в средней части соединены между собой, а спереди и сзади расходятся, образуя столбы и ножки свода. Спереди от столбов свода находится передняя спайка. Между мозолистым телом и сводом натянута тонкая вертикальная пластинка мозговой ткани – прозрачная перегородка.
Полушария имеют верхнелатеральную, медиальную и нижнюю поверхности. Верхнелатеральная – выпуклая, медиальная – плоская, обращенная к такой же поверхности другого полушария, и нижняя – неправильной формы. На трех поверхностях располагаются глубокие и мелкие борозды, и между ними извилины. Борозды – углубления между извилинами. Извилины – возвышения мозгового вещества.
Поверхности полушарий большого мозга отделены друг от друга краями – верхним, нижнелатеральным и нижневертикальным. В пространстве между двумя полушариями входит серп большого мозга – большой серповидный отросток, представляющий собой тонкую пластинку твердой оболочки, которая проникает в продольную щель большого мозга, не достигая мозолистого тела, и отделяет друг от друга правое и левое полушария. Наиболее выступающие участки полушарий получили название полюсов: лобного, затылочного и височного. Рельеф поверхностей полушарий большого мозга очень сложен и связи с наличием более или менее глубоких борозд большого мозга и расположенных между ними валикообразных возвышений – извилин. Глубина, протяженность некоторых борозд и извилин, их форма и направление очень изменчивы.
Каждое полушарие делится на доли – лобную, теменную, затылочную, височную, островковую. Центральная борозда (Роландова борозда) отделяет лобную долю от теменной, латеральная борозда (Сильвиева борозда) – височную от лобной и теменной, теменно-затылочная разделяет теменную и затылочную доли. Латеральная борозда закладывается к 4-му месяцу внутриутробного развития, теменно-зытылочная и центральная – к 6-му месяцу. Во внутриутробном периоде происходит гирификация – формирование извилин. Эти три борозды возникают первыми и отличаются большой глубиной. Вскоре к центральной борозде прибавляется еще пара ей параллельных: одна проходит впереди центральной и соответственно называется предцентральной, которая распадается на две – верхнюю и нижнюю. Другая борозда располагается позади центральной и называется постцентральной.
Постцентральная борозда лежит позади центральной борозды и почти параллельно ей. Между центральной и постцентральной бороздами располагается постцентральная извилина. Вверху она переходит на медиальную поверхность полушария большого мозга, где соединяется с предцентральной извилиной лобной доли, образуя вместе с нею парацентральную дольку. На верхнелатеральной поверхности полушария, внизу постцентральная извилина также переходит в предцентральную извилину, охватывая снизу центральную борозду. Она параллельна верхнему краю полушария. Кверху от внутритеменной борозды находится группа мелких извилин, получивших название верхней теменной дольки. Ниже этой борозды лежит нижняя теменная долька, в пределах которой выделяют две извилины: надкраевую и угловую. Надкраевая извилина охватывает конец латеральной борозды, а угловая – конец верхней височной борозды. Нижняя часть нижней теменной дольки и прилежащие к ней нижние отделы постцентральной извилины месте с нижней частью предцентральной извилины, нависающие над островковой долей, образуют лобно-теменную покрышку островка.
Доли мозга – дорсальную и латеральную поверхность коры головного мозга – принято делить на четыре доли, которые получили наименование от соответствующих костей черепа: лобная, теменная, затылочная, височная.
Затылочная доля располагается позади теменно-затылочной борозды и ее условного продолжения на верхнелатеральной поверхности полушария. По сравнению с другими долями она имеет небольшие размеры. Кзади затылочная доля кончается затылочным полюсом. Борозды и извилины на верхнелатеральной поверхности затылочной доли очень вариабельны. Наиболее часто и лучше других выражена поперечная затылочная борозда которая является как бы продолжением кзади внутритеменной борозды теменной доли мозга.
Височная доля занимает нижнебоковые отделы полушария и отделяется от лобной и теменной долей глубокой латеральной бороздой. Край височной доли, прикрывающий островковую долю, получил название височной покрышки островка. Передняя часть височной доли образует височный полюс. На боковой поверхности височной доли видны две борозды: верхняя и нижняя височные, почти параллельные латеральной борозде. Извилины височной доли ориентированы вдоль борозд. Верхняя височная извилина расположена между латеральной бороздой вверху и верхней височной внизу. На верхней поверхности этой извилины, скрытой в глубине латеральной борозды, располагаются 2-3 короткие поперечные височные извилины (извилины Гешля), разделенные поперечными височными бороздами. Между верхней и нижней височными бороздами находится средняя височная извилина. Нижнелатеральный край височной доли занимает нижняя височная извилина ограниченная сверху одноименной бороздой. Задний конец этой извилины продолжается в затылочную долю.
Островковая доля (островок) находится в глубине латеральной борозды. Эту долю можно увидеть, если раздвинуть или удалить прикрывающие островок участки: лобной, теменной и височной долей, которые получили наименование покрышки. Глубокая круговая борозда островка отделяет островок от окружающих его отделов мозга. Поверхность островка представлена длинной и короткими извилинами. Между длинной извилиной, находящейся в задней части островка и ориентированной сверху вниз и вперед, и короткими извилинами, занимающими верхнюю часть островка, находится центральная борозда островка. Нижнепередняя часть островка лишена борозд, имеет небольшое утолщение, получившее название «порог островка».
Медиальная поверхность полушария. Все доли полушария, за исключением островковой, принимают участие в образовании его медиальной поверхности (Рис. 5).

Над мозолистым телом, отделяя его от остальных отделов полушария, находится борозда мозолистого тела. Огибая сзади валик мозолистого тела, она направляется книзу и вперед и продолжается в борозду гиппокампа или гиппокампальную борозду. Выше борозды мозолистого тела находится поясная борозда. Эта борозда начинается кпереди и книзу от клюва мозолистого тела, поднимается вверх, затем поворачивает назад и следует параллельно борозде мозолистого тела, заканчиваясь выше и кзади от валика мозолистого тела под названием подтеменной борозды. На уровне валика мозолистого тела от поясной борозды вверх ответвляется краевая часть, уходящая вверх и кзади к верхнему краю полушария большого мозга. Между бороздой мозолистого тела и поясной бороздой находится поясная извилина, охватывающая мозолистое тело спереди, сверху и сзади. Сзади и книзу от валика мозолистого тела поясная извилина сужается, образуя перешеек поясной извилины. Далее книзу и кпереди перешеек переходит в более широкую парагиппокампальную извилину, ограниченную сверху бороздой гиппокампа. Поясная извилина, перешеек и парагиппокампальная извилина известны как сводчатая извилина. В глубине борозды гиппокампа находится довольно тонкая полоска серого цвета, разделенная мелкими поперечными бороздками, – зубчатая извилина. Участок медиальной поверхности полушария, находящийся между поясной бороздой и верхним краем полушария, относится к лобной и теменной долям.
Кпереди от верхнего края центральной борозды находится медиальная поверхность верхней лобной извилины, а непосредственно к указанному участку центральной борозды прилегает парацентральная долька, ограниченная сзади краевой частью поясной борозды. Между краевой частью спереди и теменно-затылочной бороздой сзади находится предклинье – принадлежащий теменной доле участок полушария большого мозга.
На медиальной поверхности затылочной доли расположены сливающиеся друг с другом под острым углом, открытым кзади, две глубокие борозды. Это теменно-затылочная борозда, отделяющая теменную долю от затылочной, и шпорная борозда, начинающаяся на медиальной поверхности затылочного полюса и направляющаяся вперед до перешейка поясной извилины. Участок затылочной доли, лежащий между теменно-затылочной и шпорной бороздами и имеющий форму треугольника, обращенного вершиной к месту слияния этих борозд, называется «клином». Хорошо заметная на медиальной поверхности полушария шпорная борозда ограничивает сверху язычную извилину, простирающуюся от затылочного полюса сзади до нижней части перешейка поясной извилины. Снизу от язычной извилины располагается коллатеральная борозда, принадлежащая уже нижней поверхности полушария.
Нижняя поверхность полушария. Рельеф нижней поверхности полушария очень сложен (Рис. 6). Передние отделы нижней поверхности образованы лобной долей полушария, позади которой выступает височный полюс, а также находятся нижние поверхности височной и затылочной долей, переходящие одна в другую без заметных границ.

На нижней поверхности лобной доли, несколько латеральнее и параллельно продольной щели большого мозга, находится обонятельная борозда. Снизу к ней прилегают обонятельная луковица и обонятельный тракт, переходящий сзади в обонятельный треугольник, в области которого видны медиальная и латеральная обонятельные полоски. Участок лобной доли между продольной щелью большого мозга и обонятельной бороздой получил название прямой извилины. Поверхность лобной доли, лежащая латеральнее от обонятельной борозды, разделена неглубокими глазничными бороздами на несколько вариабельных по форме, расположению и размерам глазничных извилин.
В заднем отделе нижней поверхности полушария хорошо видна коллатеральная борозда, лежащая книзу и латерально от язычной извилины на нижней поверхности затылочной и височной долей, латерально от парагиппокампальной извилины. Несколько кпереди от переднего конца коллатеральной борозды находится носовая борозда, ограничивающая с латеральной стороны изогнутый конец парагиппокампальной извилины – крючок. Латеральнее коллатеральной борозды лежит медиальная затылочно-височная извилина.
Между этой извилиной и расположенной кнаружи от нее латеральной затылочно-височной извилиной находится затылочно-височная борозда. Границей между латеральной затылочно-височной и нижней височной извилинами служит не борозда, а нижнелатеральный край полушария большого мозга.
Верхнелатеральная поверхность полушария – находящаяся в переднем отделе каждого полушария большого мозга лобная доля, оканчивающаяся спереди лобным полюсом и ограничивающая снизу латеральной (сильвиевой) бороздой, а сзади – глубокой центральной бороздой.
Ряд отделов головного мозга, расположенных преимущественно на медиальной поверхности полушария и являющихся субстратом для формирования таких общих состояний, как бодрствование, сон, эмоции и др., выделяют под названием «лимбическая система». Поскольку эти реакции сформировались в связи с первичными функциями обоняния (в филогенезе), их морфологической основой являются отделы мозга, которые развиваются из нижних отделов мозгового пузыря и относятся к так называемому обонятельному мозгу. Лимбическую систему составляют обонятельная луковица, обонятельный тракт, обонятельный треугольник, переднее продырявленное вещество, которые расположены на нижней поверхности лобной доли (периферический отдел обонятельного мозга), а также поясная и парагиппокампальная (вместе с крючком) извилины, зубчатая извилина, гиппокамп (центральный отдел обонятельного мозга) и некоторые другие структуры. Включение этих отделов мозга в лимбическую систему оказалось возможным в связи с общими чертами их строения (и происхождения), наличием взаимных связей и сходством функциональных реакций.
Полушария состоят из серого и белого вещества. Слой серого вещества называется корой головного мозга. Кора покрывает в виде плаща остальные образования большого мозга и поэтому и называется плащом. Под корой белое вещество, а в нем островки серого вещества – базальные ядра, в основном расположенные в лобной доле. К ним относят полосатое тело (хвостатое ядро и чечевицеобразное ядро), ограду и миндалевидное тело.
Полосатое тело (стриопаллидарная система) состоит из 2-х ядер – хвостатого и чечевицеобразного, разделенных прослойкой белого вещества – внутренней капсулой. В эмбриональном периоде полосатое тело составляет одну серую массу, затем оно разделяется. Хвостатое ядро расположено около таламуса. Оно имеет подковообразную форму, состоит из головки, тела и хвоста. Чечевицеобразное ядро имеет форму чечевичного зерна, находится латеральнее таламуса и хвостатого ядра. Чечевицеобразное ядро делится на 3 части, благодаря белому веществу. Наиболее латерально лежит скорлупа, имеющая темную окраску, а две более светлые части называются латеральным и медиальным бледными шарами.
Для скорлупы характерно участие в организации пищевого поведения: пищепоиска, пищенаправленности, пищезахвата и пищевладения; ряд трофических нарушений кожи, внутренних органов возникает при нарушениях функции скорлупы. Раздражения скорлупы приводят к изменениям дыхания, слюноотделения.
Бледный шар имеет преимущественно крупные нейроны Гольджи 1-го типа. Связи бледного шара с таламусом, скорлупой, хвостатым ядром, средним мозгом, гипоталамусом, соматосенсорной системой и др. свидетельствуют о его участии в организации простых и сложных форм поведения.
Раздражение бледного шара с помощью вживленных электродов вызывает сокращение мышц конечностей, активацию или торможение γ-мотонейронов спинного мозга. У больных с гиперкинезами раздражение разных отделов бледного шара (в зависимости от места и частоты раздражения) увеличивает или снижает гиперкинез.
Стимуляция бледного шара, в отличие от стимуляции хвостатого ядра, не вызывает торможения, а провоцирует ориентировочную реакцию, движения конечностей, пищевое поведение (обнюхивание, жевание, глотание и т.д.).
Повреждение бледного шара вызывает у людей гипомимию, маскообразность лица, тремор головы, конечностей (причем этот тремор исчезает в покое, во сне и усиливается при движениях), монотонность речи. При повреждении бледного шара наблюдается миоклония – быстрые подергивания мышц отдельных групп или отдельных мышц рук, спины, лица.
Ядра полосатого тела конечного мозга являются подкорковыми двигательными центрами в составе экстрапирамидной системы, регулирующими сложные автоматизированные двигательные акты. К экстрапирамидной системе относят черное вещество и красные ядра ножек мозга. Полосатое тело управляет процессами теплорегуляции и обмена углеводов. К наружи от чечевицеобразного ядра расположена тонкая пластинка серого вещества – ограда.
Ограда находится в белом веществе полушария сбоку от скорлупы, между последней и корой островковой доли. Она содержит полиморфные нейроны разных типов и образует связи преимущественно с корой большого мозга. Глубокая локализация и малые размеры ограды представляют определенные трудности для ее физиологического исследования.
Миндалевидное тело (большая спайка мозга) находится в переднем отделе височной доли, входит в состав лимбической системы. К белому веществу полушария относятся внутренняя капсула и волокна, проходящие спайки (мозолистое тело, передняя спайка, спайка свода) и направляющиеся к коре и базальным ядрам.
Внутренняя капсула – толстая изогнутая пластинка белого вещества. Она делится на три отдела: 1) переднюю ножку внутренней капсулы, 2) заднюю ножку внутренней капсулы, 3) место соединения этих двух отделов – колено внутренней капсулы. В колене внутренней капсулы располагаются корково-ядерные пути, идущие к двигательным ядрам черепных нервов. В переднем отделе располагаются корково-спинномозговые волокна, находящиеся в предцентральной извилине и идущие к двигательным ядрам передних рогов спинного мозга. В задней ножке располагаются таламокортикальные волокна, идущие в кору постцентральной извилины. В состав этого проводящего пути соединяются волокна проводников всех видов общей чувствительности (большой температуры, осязания, давления, проприоцентивной). В задних отделах задней ножки располагаются слуховой и зрительный проводящие пути. Оба берут начало от подкорковых центров слуха и зрения и заканчиваются в соответствующих центрах.
Таким образом, базальные ядра головного мозга являются интегративными центрами организации моторики, эмоций, высшей нервной деятельности, причем каждая из этих функций может быть усилена или заторможена активацией отдельных образований базальных ядер.
Мозолистое тело представляет собой толстую изогнутую пластинку, состоящую из поперечных волокон.
В мозолистом теле разделяют: колено, клюв, между ними ствол, который переходит в валик. Волокна, проходящие в колене, соединяют кору лобных долей правого и левого полушарий. Волокна ствола соединяют серое вещество теменных и височных долей. В валике проходящие волокна соединяют кору затылочных долей.
Под мозолистым телом располагается свод, который состоит из двух дугообразно изогнутых тяжей, соединенных при помощи спайки. Свод состоит из тела, парного столба и парных ножек. Ножки, срастаясь с гипокампом, образуют бахромку.
Боковой желудочек – полость полушарий (I и II желудочки) и сообщающяеся через межжелудочковое отверстие с III желудочком. В каждом желудочке выделяют центральную часть и три рога: передний (лобный) – в лобной доле; задний (затылочный) – в затылочной доле и нижний (височный) – в височный доле. Боковые желудочки, как и другие желудочки головного мозга, и центральный канал спинного мозга изнутри выстланы слоем эпендимоцитов. Эпендимные клетки образуют спинномозговую жидкость и регулируют ее состав.
Ромбовидная ямка представляет собой ромбовидной формы вдавление, длинная ось которой направлена вдоль мозга. Она ограничена с боков в своем верхнем отделе верхними мозжечковыми, в нижнем – нижними мозжечковыми ножками.
ПОЛУШАРИЯ БОЛЬШОГО МОЗГА — Мегаобучалка
ГОУ СПО «Кемеровский Областной Медицинский Колледж»
Реферат по анатомии и физиологии человека
Тема: Головной мозг человека
Выполнил студент : Привалихина Анна
Группа МС 103Д
Проверил преподаватель: Шарыгина Елена Олеговна
Кемерово, 2011
План
Головной мозг человека
Полушария большого мозга
Продолговатый мозг
Средний мозг
Промежуточный мозг
Мост
Мозжечок
Кора большого мозга
Оболочки головного мозга
Боковые желудочки
1 — обонятельная луковица;
2 — обонятельный тракт;
3 — переднее продырявленное вещество;
4 — серый бугор;
5 — зрительный тракт;
6- сосцевидные тела;
7 — троичный узел;
8 — заднее продырявленное вещество;
9 — мост;
10 -мозжечок;
11 — пирамида продолговатого мозга;
12 — олива;
13 — спинномозговые нервы;
14 — подъязычный нерв;
15 — добавочный нерв;
16 — блуждающий нерв;
17 — языкоглоточный нерв;
18 — преддверно -улитковый нерв;
19 — лицевой нерв;
20 — отводящий нерв;
21 — троичный нерв;
22 — боковой нерв;
23 — глазодвигательный нерв;
24 — зрительный нерв;
25 -обонятельные нервы.
ГОЛОВНОЙ МОЗГ
Головной мозг, с окружающими его оболочками находится в полости мозгового черепа. Верхняя вентральная поверхность головного мозга по форме соответствует внутренней вогнутой поверхности свода черепа. Нижняя поверхность — основание головного мозга, имеет сложный рельеф, соответствующий черепным ямкам внутреннего основания черепа.
Масса мозга взрослого человека колеблется от 1100 до 2000 г. На протяжении от 20 до 60 лет масса и объем остаются максимальным и постоянным для каждого индивидуума.
ПОЛУШАРИЯ БОЛЬШОГО МОЗГА
У взрослого человека — это наиболее сильно развитая, самая крупная и функционально наиболее важная часть ЦНС. Отделы полушарий прикрывают собой все остальные части головного мозга. Правое и левое полушария отделены друг от друга глубокой продольной щелью большого мозга, достигающий большой спайки мозга, или мозолистого тела.
В задних отделах продольная щель впадает в поперечную щель большого мозга, которая отделяет полушария от мозжечка.
На вентральной, медиальной и нижней поверхностях полушарий головного мозга расположены глубокие и мелкие борозды. Глубокие борозды разделяют каждое из полушарий на доли большого мозга. Мелкие борозды отделяют друг от друга извилины большого мозга. Нижняя поверхность, или основание, головного мозга образована вентральными поверхностями полушарий большого мозга, мозжечка и вентральными отделами мозгового ствола.
К задней поверхности зрительного перекреста прилежит серый бугор, нижние отделы которого вытянуты в виде постепенно суживающейся к низу трубки — воронки. На нижнем конце воронки располагается округлое образование — гипофиз. К серому бугру примыкают два белых шарообразных возвышения — сосцевидных тела.
Сзади от зрительных трактов видны два продольных белых валика — ножки мозга, между которыми находится углубление — межножковая ямка. Дно ее образовано задним продырявленным веществом. Еще дальше располагается широкий поперечный валик — мост. Латеральные отделы моста продолжаются в мозжечок, образуя его средние мозжечковые ножки.
Каудальнее моста отделы продолговатого мозга представлены медиально расположенными пирамидами, разделенными друг от друга передней серединной щелью, а латерально — оливами.
Осмотр медиальной поверхности полушарий большого мозга , некоторых деталей мозгового ствола и мозжечка становиться возможным при проведении серединного разреза по продольной щели большого мозга .
Обширная медиальная поверхность полушарий большого мозга нависает над значительно меньшими по размерам мозжечком и мозговым стволом. На медиальной поверхности полушарий, как и на других поверхностях, видны борозды, которые отделяют друг от друга извилины.
Участки лобной, теменной и затылочной долей отделены от мозолистого тела одноименной бороздой.
Серединная часть мозолистого тела носит название ствола. Передние отделы его загибаются к низу, образуя колено мозолистого тела. Еще более книзу мозолистое тело истончается и переходит в клюв мозолистого тела. Задние отделы мозолистого тела в средней его части отделяется тонкая белая пластинка, называемая телом свода. Постепенно отделяясь от мозолистого тела и образуя дугообразный изгиб вперед и книзу, тело продолжается в столб свода, который заканчивается сосцевиднымм телом , сзади — в ножки свода. Между столбами свода поперечно проходит пучок нервных волокон, заметный на срезе в виде белого овала, — это передняя спайка мозга. Как и поперечно идущие волокна мозолистого тела, они связывают друг с другом полушария большого мозга.
Столбы свода окружают тонкую пластинку мозгового вещества — прозрачную перегородку.
Все перечисленные образования головного мозга относятся к конечному мозгу.
Структуры, расположенные ниже, за исключением мозжечка, относятся к мозговому стволу (промежуточный, средний, задний отделы головного мозга и продолговатый мозг).
Самые передние отделы мозгового ствола образованы зрительными буграми, которые расположены к низу от тела свода и мозолистого тела и позади столбов свода.
На срединном разрезе мозга видна только медиальная поверхность заднего таламуса (зрительного бугра).
В задневерхних отделах зрительных бугров находится шишковидное тело, передненижние отделы которого срастаются тонким поперечно идущим тяжем ( задняя спайка).
Зрительные бугры и расположенные рядом с ним образования, описанные выше, относятся к промежуточному мозгу.
К задней поверхности зрительного бугра примыкают образования, относящиеся к среднему мозгу.
ПРОДОЛГОВАТЫЙ МОЗГ
Является непосредственным продолжением спинного мозга. Граница между продолговатым и спинным мозгом соответствует уровню краев большого затылочного отверстия. Верхняя граница продолговатого мозга на вентральной поверхности проходит по заднему краю моста.
Передние отделы продолговатого мозга по сравнению с задними несколько утолщаются и этот отдел мозга приобретает форму усеченного конуса. Борозды продолговатого мозга являются продолжением борозд спинного мозга и носят те же названия. По обеим сторонам от передней срединной щели на вентральной поверхности продолговатого мозга расположены выпуклые, постепенно суживающиеся к низу пирамиды.
Латеральнее пирамиды с обеих сторон находятся овальные возвышения — оливы.
В нижней части на дорсальной поверхности продолговатого мозга тянется задняя серединная борозда, по бока от которой заканчивается утолщениями тонкий и клиновидный пучки задних канатиков спинного мозга. В этих утолщениях располагаются ядра этих пучков, от которых отходят волокна, формирующие медиальную петлю. Медиальная петля на уровне продолговатого мозга образует перекрест. Пучки этого прекреста расположены дорсальнее пирамид, в межоливном слое. Здесь же проходят волокна медиального пучка. Латеральнее оливы из задней латеральной борозды выходят тонкие корешки языкоглоточного, блуждающего и добавочных нервов, ядра которых лежат в дорсолатеральных отделах продолговатого мозга.
Серое вещество продолговатого мозга представлено в вентральных отделах скоплениями нейронов, которые образуют нижние оливные ядра. Дорсальнее пирамид вдоль всего продолговатого мозга располагается ретикулярная формация, которая представлена переплетением нервных волокон и лежащими между ними нервными клетками.
На уровне продолговатого мозга находятся такие жизненно важные центры, как дыхательный и кровообращения.
СРЕДНИЙ МОЗГ
Состоит из дорсального отдела крыши среднего мозга и вентрального — ножек мозга, которые разграничиваются полостью — водопроводом мозга. Нижней границей среднего мозга на его вентральной поверхности является передний край моста, верхний зрительный тракт и уровень сосцевидных тел. На препарате головного мозга пластинку четверхоломия, или крышу среднего мозга, можно увидеть лишь после удаления полушарий большого мозга.
На основании головного мозга хорошо видна вторая часть среднего мозга в виде двух толстых белых расходящихся пучков, идущих в ткань полушарий большого мозга, — это ножки мозга. Углубление между правой и левой ножками мозга называются межножковой ямкой, из нее выходят корешки глазодвигательных нервов. Впереди о ядра глазодвигательного нерва лежит ядро медиального продольного пучка. Самым крупным ядром среднего мозга является красное ядро — одно из центральных координационных ядер экстрапирамидной системы. Рядом с водопроводом лежит ретикулярная форма среднего мозга.
На поперечном разрезе отчетливо видно черепное вещество, которое делит ножку мозга на два отдела: дорсальный — покрышку среднего мозга и вентральный — основание ножки мозга. В покрышке среднего мозга располагаются ядра среднего мозга и проходят восходящие проводящие пути. Вентральные отделы ножек мозга целиком состоят из белого вещества, здесь проходят нисходящие проводящие пути.
Функциональное значение среднего мозга состоит в том. что здесь расположены подкорковые центры слуха и зрения; ядра головных нервов, обеспечивающих иннервацию поперечнополосатых и гладких мышц глазного яблока: ядра, относящиеся к экстрапирамидной системе, обеспечивающей сокращение мышц тела во время автоматических движений. Через средний мозг следуют нисходящие (двигательные) и восходящие (чувствительные) проводящие пути. Область среднего мозга является также местом расположения вегетативных центров (центральное серое вещество) и ретикулярной формации.
ПРОМЕЖУТОЧНЫЙ МОЗГ
Представлен следующими отделами
1) областью зрительных бугров (таламическая область), которая расположена в дорсальных его участках;
2) гипоталамусом (подталамическая область), составляющим вентральные отделы промежуточного мозга;
3) III желудочком, имеющим вид продольной (сагитальной) щели между правым и левым зрительными буграми и соединяющимися через межжелудочковое отверстие с боковыми желудочками.
В свою очередь таламическая область подразделяется на таламус (зрительный бугор), метаталамус (медеальное и латеральное коленчатые тела) и эпиталамус (шишковидное тело, поводки, спайки поводков и эпиталамическая спайка).
Зрительные бугры состоят из серого вещества, в котором различают отдельные скопления нервных клеток (ядра зрительного бугра), разделенными тонкими прослойками белого вещества. В связи с тем что здесь переключается большая часть чувствительных проводящих путей, зрительный бугор фактически является подкорковым чувствительным центром, а его подушка — подкорковым зрительным центром.
К медиальной поверхности зрительных бугров при помощи поводков присоединяется шишковидное тело — эпифиз.
Гипоталамус составляет вентральный отдел промежуточного мозга и участвует в образовании дна III желудочка. К гипоталамусу относятся серый бугор с воронкой и гипофизом — железной внутренней секреции, зрительный тракт, зрительный перекрест, сосцевидные тела.
Гипоталамус представляет собой продолжение ножек мозга в промежуточный мозг. Серое вещество подталамической области располагается в виде ядер, способных вырабатывать нейросекрет и транспортировать его в гипофиз, регулируя эндокринную работу последнего.
Таким образом, серое вещество промежуточного мозга составляют ядра, относящиеся к подкорковым центрам всех видов чувствительности. В области промежуточного мозга расположены ретикулярная формация, центры экстрапирамидной системы, вегетативные центры, регулирующие все виды обмена веществ и нейросекретные ядра.
Белое вещество промежуточного мозга представлено проводящими путями восходящего и нисходящего направлений, обеспечивающих двустороннюю связь коры головного мозга с подкорковыми образованиями и центрами спинного мозга. Помимо этого, к промежуточному мозгу относятся две железы внутренней секреции — гипофиз и шишковидное тело, принимающие участие вместе с соответствующими ядрами гипоталамуса и эпиталамуса и образовании гипоталамогипофмзарной и эпиталамо-эпифизарной систем.
КОНЕЧНЫЙ МОЗГ
Состоит из двух полушарий большого мозга, каждое из которых представлено плащом, обонятельным мозгом и базальными ядрами. Полостью конечного мозга являются боковые желудочки, находящиеся в каждом из полушарий. Полушария большого мозга отделены друг от друга продольной щелью большого мозга и соединяются при помощи мозолистого тела , передней и задней спаек и спайки свода. Мозолистое тело состоит из поперечных волокон которые в латеральном направлении продолжаются в полушария, образуя лучистость мозолистого тела, соединяя друг с другом участки лобных и затылочных долей полушарий, дугообразно изгибаются и образуют передние — лобные и задние — затылочные щипцы. К задней и средней частям мозолистого тела снизу прилежит свод мозга, состоящий из двух дугообразно изогнутых тяжей, сращенных в средней своей части при помощи переднее спайки мозга.
МОСТ
На основании мозгового ствола имеет вид поперечно расположенного белого валика, который имеет вид поперечно расположенного белого валика, который в каудальном отделе граничит с пирамидами и оливами продолговатого мозга, а в краниальном — с ножками мозга.
Продолжение моста в латеральном направлении образует среднюю ножку мозжечка.
Дорсальная поверхность моста прикрыта мозжечком и снаружи не видна.
На поперечном разрезе моста в центральных отделах можно видеть толстый пучок идущих поперечно волокон, относящихся к проводящему пути слухового анализатора и образующих трапецивидное тело.
В нижних отделах моста заметны скопления серого вещества, называемые ядрами собственно моста, которые выступают в роли посредников в осуществлении связей коры полушарий большого мозга с полушариями мозжечка.
В дорсальной части моста лежат волокна медиальной петли, идущей от продолговатого мозга. над которым расположена ретикулярная формация моста. Латеральнее проходят волокна слуховой петли.
МОЗЖЕЧОК
Составляет более крупную, чем мост, часть заднего мозга, которая заполняет собой большую часть задней черепной ямки.
В мозжечке различают верхнюю и нижнюю поверхности, границами между которыми являются передний и задний края.
Верхняя поверхность мозжечка на целом мозге прикрыта затылочными долями полушарий большого мозга и отделена от них глубокой поперечной щелью большого мозга. В мозжечке различают непарную серединную часть — червь, два полушария. поперечными бороздами червь расчленен на мелкие извилины, которые придают ему некоторое сходство с кольчатым червем. Обе поверхности полушарий и червя изрезанны множеством поперечных параллельно идущих мелких бороздок, между которыми находятся длинные и узкие извилины мозжечка. Группа извилин, отделенных более глубокими бороздами, образуют дольки мозжечка. Полушария мозжечка и червь состоят из белого вещества, расположенного внутри, и тонкой прослойки серого вещества коры мозжечка, окаймляющего белое вещество по периферии. Кора мозжечка представлена тремя слоями нервных клеток. На сагитальном разрезе белое вещество мозжечка представлена тремя слоями нервных клеток и имеет вид ветвистого дерева.
В толще белого вещества обнаруживаются отдельные парные скопления нервных клеток, которые образуют зубчатое, пробковидное, шаровидное ядра мозжечка и ядра шатра.
В мозговом стволе следующим после моста отделом, небольшим , но функционально важным , является перешее ромбовидного мозга, состоящий из верхних ножек мозжечка, верхнего мозгового паруса и треугольной петли, в котором проходят волокна латеральной (слуховой ) петли.
КОРА БОЛЬШОГО МОЗГА
Образована белым и серым веществом. В коре выделяют 6 слоев нервных клеток, различные ее отделы имеют разную толщину (от 1,5 до 5,0 мм, в среднем 2-3 мм). Каждое из полушарий имеет три поверхности : наиболее выпуклую верхнелатеральную, плоскую, обращенную к противоположному полушарию медиальную и имеющую сложный рельеф, соответствующий внутреннему основанию черепа, — нижнюю, поверхность полушария или основание мозга. Наиболее выступающие участки полушарий получили название лобного, затылочного, височного полюсов. Поверхность полушарий изрезанна глубокими щелями, бороздами. Усложняют рельеф расположенные между ними участки — извилины. Глубина, протяженность борозд, их форма и направление очень изменчивы.
Щели и борозды подразделяют полушария на лобную, теменную, височную, затылочную и островковую доли. Последняя не видна при обзоре поверхностей полушарий, т.к. островок находится на дне латеральной борозды и прикрыт участками других долей.
На верхенлатеральной поверхности полушария обращает на себя внимание латеральная борозда, которая является границей между лобной, теменной и височными долями и идет от нижней поверхности полушарий назад и вверх.
Другая крупная борозда — центральная борозда. Начинается приблизительно от середины верхнего края полушарий и следует вниз и несколько вперед, но не достигает латеральной борозды. Центральная борозда отделяет лобную долю от теменной. Выраженная граница между темеенной и затылочными долями на дорсолатеральной поверхности полушарий отсутствует.
Лобная доля. Впереди от центральной борозды почти параллельно ей тянется предцентральная борозда, которая дает начало двум параллельным бороздам, идущим к лобному полюсу. Названные борозды делят поверхность мозга на лежащую перед центральной бороздой предцентральную извилину и горизонтально идущие верхнюю, среднюю и нижнюю лобные извилины.
Теменная доля. Сзади от центральной борозды и почти параллельно ей проходит постцентральная борозда, от которой в сторону затылочной доли направляется продольная внутритеменная борозда. Эти две борозды делят теменную долю на постцентральную извилину, а также на верхнюю и нижнюю теменные дольки.
Височная доля. Верхнелатеральная поверхность височной доли представлены двумя бороздами, идущими параллельно латеральной борозде, которые делят поверхность мозга на верхнюю , среднюю и нижнюю извилины.
Серое вещество полушарий большого мозга представлено корой и базальными ядрами конечного мозга. К базальным ядрам относятся полосатое тело, состоящее из хвостатого и чечевицеобразного ядер; ограда и миндалевидное тело. Прослойки белого вещества между ними образуют наружную и внутреннюю капсулы, причем последняя представляет собой толстый слой белого вещества, состоящий из проводящих путей головного мозга. Во внутренней капсуле и выделяют переднюю и заднюю ножки и колено.
Борозды и извилины: порерхность коры головного мозга
Характерная особенность человеческого мозга – невероятный размер коры и сложная складчатость. Кора – наиболее развитая область головного мозга, отвечающая за нерефлекторную деятельность (память, восприятие, познание, мышление и т. п.).
Формирование корково-подкорковых структур происходит во время эмбрионального развития, обеспечивая возможность размещения коры в ограниченном объеме черепной коробки. Извилины (гири) и борозды (сульчи) составляют её сложенную поверхность. Патологические изменения в размерах или складках коры приводят к тяжелой умственной неполноценности и трудноизлечимой эпилепсии. Следовательно, корковое расширение и складчатость рассматриваются как ключевые процессы в эволюции мозга.
Борозды и извилины: формирование и функции
Борозды и извилины в нейроанатомии, придающие головному мозгу морщинистый вид, служат двум важнейшим функциям. Они помогают увеличить площадь поверхности коры, что позволяет большему количеству нейронам уплотняться в неё и усиливать способность мозга обрабатывать информацию. Борозды и извилины головного мозга образуют деления, создавая границы между долями мозга, разделяя его на два полушария.

Основные борозды:
- Межполушарная щель – глубокая борозда по центру мозга, в которой находится мозолистое тело.
- Сильвиевая щель (латеральная борозда) разделяет теменную и лобную доли.
- Роландова щель (центральная борозда), отделяющая веретенообразную извилину и извилину гиппокампа на нижней поверхности височных долей.
- Теменно-затылочная — разделяет теменную и затылочную доли.
- Шпорная щель (шпороподобная борозда или выделяющаяся фиссура) – расположена в затылочных долях, делит зрительную кору.
Основные извилины мозга:
- Ангулярная извилина теменной доли помогает в обработке слухового и визуального распознавания.
- Извилина Брока (центр Брока) – область мозга, расположенная в левой лобной доле у большинства людей, которая контролирует функции, связанные с воспроизведением речи.
- Поясная извилина – арочная складка, расположенная над мозолистым телом, является компонентом лимбической системы и обрабатывает сенсорный ввод относительно эмоций, регулирует агрессивное поведение.
- Веретенообразная извилина находится в височной и затылочной долях и состоит из боковых и медиальных частей. Считается, что играет роль в распознавании слов и лиц.
- Гиппокампальная извилина складывается на внутренней поверхности височной доли, которая граничит с гиппокампом. Играет важную роль для памяти.
- Язычная извилина в затылочной доле, участвующая в зрительной обработке. Она ограничена коллатеральной бороздой и шпорной щелью. Спереди соприкасается с парарпопампальной извилиной, и вместе они образуют медиальную часть веретенообразной извилины.
По мере развития эмбриона извилины и борозды формируются с появлением углублений на поверхности коры. Не все извилины развиваются одновременно. Первичная форма образуется, начиная с 10 недели беременности (у человека), затем развиваются вторичные и третичные. Наиболее выдающаяся борозда – латеральная. За ней следует центральная, отделяющая моторную кору (прецентральную извилину) от соматосенсорной коры (постцентральной извилины). Большинство кортикальных борозд и извилин головного мозга, анатомия которых начинает складываться между 24 и 38 неделями беременности, продолжают расти и развиваться после того, как новорожденный появится на свет.
Раннее состояние головного мозга оказывает сильное влияние на конечный уровень гирификации. В частности, существует обратная связь между корковой толщиной и гирификацией. Участки мозга с низким значением толщины имеют более высокий уровень гирификации. Также верно и обратное, что участки мозга с высоким значением толщины (например, утолщение коры гиппокампальных извилин головного мозга) – низкий уровень гирификации.
Доли мозга и их функции
Каждое из полушарий разделено на четыре доли: лобную, теменную, височную и затылочную. Большинство функций мозга полагаются на разные области по всему мозгу, которые работают вместе, но каждая доля выполняется основную часть относительно определенных функций.

Лобная доля расположена в самой передней области коры головного мозга, отделена от теменной доли центральной бороздой, от височной – боковой. В области, как правило, сосредоточены наиболее значимые для человека исполнительные функции, включая регулирование эмоций, планирование, рассуждение, решение проблем.
Теменная доля ответственна за интеграцию сенсорной информации, включая контакт, температуру, давление, боль. Из-за обработки, возникающей в париетальной доле, возможно различать прикосновение двух объектов в близлежащих точках (а не как один объект). Этот процесс называют двухточечным.
Височная доля также содержит области, участвующие в обработке сенсорной информации, особенно важные для слуха, распознавания языка, формирования воспоминаний. Первичная слуховая кора получает аудио информацию через уши и вторичные области и обрабатывает данные так, чтобы человек понимал то, что он слышит (слова, смех, плач и так далее). Медиальная (ближе к центру мозга) часть содержит гиппокамп – зону, важную для памяти, обучения, восприятия эмоций. Некоторые области височной доли обрабатывают сложную визуальную информацию, включая лица и сцены.
Клеточные механизмы, приводящие к расширению и складыванию коры головного мозга
Строение мозга человека отличает его от прочих млекопитающих, и по этой причине может объяснять его уникальные умственные способности по сравнению с другими животными. Количество складок в коре, возможно, коррелирует с некоторыми специфическими когнитивными, сенсорными, двигательными способностями. Хотя нет четкого объяснения того, каким образом происходит уникальное разделение человеческого мозга на борозды и извилины. Сегодня имеется прогресс в понимании чрезвычайно сложных процессов в мозге, кора которого строится с таким количеством борозд и извилин. Несмотря на то, что у всех клеток одна и та же ДНК, образуются разные нервные стволовые клетки. Именно их работа с различными свойствами создает основную структуру мозга, состоящую из нейронов и глиальных клеток.

Теленцефалический нейроэпителий
Рост мозга происходит посредством двух видов стволовых клеток – нейронных стволовых клеток и нейронных предшественников. Обе эти формы образуют нейроны, которые становятся постоянными в мозге, а также промежуточные клетки, создающие строительный материал для построения мозга. Четыре различных типа стволовых клеток определяют строение коры.
В период раннего эмбрионального развития расширение рострального домена нервной трубки приводит к появлению двух телэнцефальных пузырей. Дорзальная половина этих пузырей молекулярно определяется как зачаток коры головного мозга. На этом этапе кортикальный зачаток состоит исключительно из монослоя нейроэпителиальных клеток-предшественников. Они сильно поляризованы и прикреплены друг к другу плотными соединениями на уровне апикального домена (внутренней поверхности телэнцефалического пузыря) и перемещают клеточное ядро между апикальной (верхушечной) и базальной (нижней) стороной нейроэпителия в согласованности с клеточным циклом.
- базально-направленное движение во время G1-фазы;
- базальное положение во время S-фазы;
- апикально-направленное движение во время G2-фазы;
- митоз на апикальной поверхности.
Циклическое движение известно как межкинетическая ядерная миграция и полностью асинхронно между нейроэпителиальными клетками, придавая нейроэпителию псевдостратированный вид. Клетки подвергаются только симметричным самоагрессирующим делениям, при этом каждое деление генерирует две дочерние клетки, следовательно, экспоненциально увеличивая их число. Поскольку они являются основополагающими клетками-предшественниками коры головного мозга, размер их объединения определяет количество производных нейрогенных клеток-предшественников и конечное число кортикальных нейронов, и, следовательно, он оказывает фундаментальное влияние на размер зрелой коры головного мозга. Увеличение количества приводит к расширению площади поверхности и формированию нейроэпителия.

Распространение и нейрогенез
Непосредственно перед началом нейрогенеза нейроэпителиальные клетки-предшественники начинают терять плотные соединения, и приобретать признаки, типичные для глиальных клеток (включая экспрессию липид-связывающего белка мозга, виментина и Pax6), становясь, таким образом, апикальными радиальными глиальными клетками (АРГК). Они также подвергаются межкинетической ядерной миграции, делятся на верхушечной поверхности развивающейся коры и на этом раннем этапе также подвергаются самоусиливающимся делениям.
Однако постепенно они начинают делиться асимметрично, чтобы генерировать одну себе подобную клетку плюс другую клетку. Эти новые клетки накапливаются в базальной части коркового зачатка, тогда как клеточные тела АРГК остаются на апикальной стороне, образуя желудочковую зону (ЖЗ). С накоплением клеток выше ЖЗ процесс АРГК продлевается, оставаясь присоединенным к базальной пластине, и теперь называется радиальной глией. Асимметричные АРГК-деления генерируют один АРГК плюс один нейрон или одну промежуточную клетку-предшественник. Промежуточные клетки-предшественники (вторичные клетки-предшественники без апикально-базальной полярности) не подвергаются межкинетической ядерной миграции, делятся в слое, расположенном в области желудочковой зоны, субвентрикулярной зоны (СВЗ), и все они выражают транскрипционный фактор (Tbr2).
Однако из-за того, что каждый нейрон сам потребляет при митозе, их относительное количество по сравнению с АРГК достаточно низкое. Промежуточные клетки-предшественники в коре головного мозга генерируют большинство кортикальных возбуждающих нейронов. По мере развития нейрогенеза потребность в расширении/обновлении АРГК снижается, и повышается в производстве нейронов. Дополнительно к расширенной желудочковой зоне утолщается субвентрикулярная зона, населенная в изобилии базальными предшественниками, особенно на поздних стадиях нейрогенеза. В результате происходит расщепление СВЗ на внутреннюю и внешнюю часть. Внешняя часть содержит большое разнообразие типов клеток-предшественников с высоким потенциалом развития, что является ключевым фактором для расширения и формирования коры.